![]()
|
|
Ustroj i djelovanje moždane kore: Uvod u neurologiju ponašanja čovjeka
doc. dr. sci. Goran Šimić,
dr. med.
Zavod za neuroznanost Hrvatskog instituta za istraživanje mozga
Medicinskog fakulteta Sveučilišta u Zagrebu
1. Uvodne napomene
2. Podjela moždane kore
2.1. Limbički korteks
2.2. Paralimbički korteks
2.3. Unimodalni i heteromodalni asocijacijski
korteks
2.4. Idiotipski korteks
3. Organizacija moždane kore i uloga transmodalnih areja u intermedijarnom
procesiranju
4. Glavne neuronske mreže moždane kore čovjeka
4.1. Mreža za prepoznavanje lica i objekata
s epicentrima u lateralnom temporalnom i temporopolarnom korteksu
4.2. Mreža prostorne pozornosti s epicentrima
u stražnjem parijetalnom korteksu, čeonom području za očne pokrete i cingularnom
girusu
4.3. Mreža jezičnih sposobnosti s epicentrima
u Wernicke i Broca području
4.4. Mreža sposobnosti pamćenja i emocija s
epicentrima u entorinalno-hipokampalnom području i amigdaloidnom kompleksu jezgara
4.5. Mreža izvršnih funkcija i kontrole načina
ponašanja s epicentrima u lateralnom prefrontalnom i orbitofrontalnom korteksu
5. Završne napomene i zaključci
Nakon što su se koncem 19. stoljeća
stvorili preduvjeti za istraživanje citoarhitektonske građe mozga (otkriće histološkog
bojanja tijela živčanih stanica po Nisslu), fine građe svih nastavaka živčanih
stanica (bojanje po Golgiju), te snopova vlakana (bojanje mijelinskih ovojnica
aksona po Weigertu), upotreba ovih metoda u usporednoj analizi živčanog sustava
kralježnjaka dovela je do zaključka da je stupanj složenosti ponašanja kralježnjaka
razmjeran stupnju razvijenosti kore (cortex) krajnjeg mozga (telencephalon).
U riba je korteks primitivan, te da se najvećim dijelom bavi jednostavnom (paleocortex)
i nešto složenijom obradom njušnih podražaja (archicortex). U ptica i vodozemaca,
naročito reptila, javlja se novi korteks (neocortex), a osim toga se i drugi
osjetni sustavi, osim njušnog, počinju značajnije projicirati u korteks, i to
preko međumozga (thalamus). U sisavaca dolazi do golemog relativnog povećanja
krajnjeg mozga u odnosu na veličinu tijela, a također i s njim povezanih subkortikalnih
struktura - talamusa i bazalnih ganglija. Ovaj proces filogenetskog povećavanja
krajnjeg mozga naziva se telencefalizacija, a najviše je izražen u čovjeka.
Ontogenetski razvitak moždane kore temelji se na migraciji neurona iz proliferativnih
zona koje oblažu neuralnu cijev. Čini se da je odvojenost proliferativnih središta
od konačnog smještaja neurona odigrala presudnu ulogu u evoluciji, jer je na
taj način bila omogućena sve složenija građa mozga. Za razliku od relativnog
kratkog razdoblja rađanja i migracije živčanih stanica, njihova maturacija je
puno dulji i složeniji proces. Rano djetinjstvo je razdoblje brzog rasta i diferencijacije
kortikalnih neurona grananjem dendrita, te porastom broja sinapsi. Tako se,
primjerice, između rođenja i druge godine života djeteta u čeonom režnju bilježi
porast broja sinapsi veći od 10 puta, a i u drugim dijelovima moždane kore je
slično. Nakon ovog početnog dijela maturacije kojim upravlja genetski program
živčanih stanica, daljnja maturacija više se ne može odvijati neovisno od utjecaja
okoline (dapače, ona mu je razmjerna). Ovi procesi stoga se nazivaju epigenetski
procesi. Takozvana 'fina' maturacija kortikalnih neuronskih krugova vjerojatno
traje do kasne adolescencije, no mogućnost daljnjeg sinaptičkog preoblikovanja
ostaje doživotno svojstvo i jedno od glavnih obilježja moždane kore, a u širem
se smislu naziva plastičnost mozga.
2. Podjela moždane kore čovjeka
Moždana kora čovjeka sadrži otprilike
20 milijardi neurona koji se rasprostiru na gotovo 2000 kvadratnih centimetara
površine. Nedostatak jasnih anatomsko-funkcionalnih granica pojedinih dijelova
moždane kore doveo je do razvitka brojnih pristupa njenoj parcelizaciji. Rezultirajuće
mape mogu se podijeliti na strukturalne, te one koje su načinjene prema pretežno
funkcionalnim obilježjima.
Od strukturalnih podjela moždane kore najčešće se koristi ona po Brodmannu,
a temelji se na citoarhitektonskim razlikama u slojevnoj (laminarnoj) građi
moždane kore (Slika 1a i 1b).
Slika 1a. Brodmannova citoarhitektonska mapa moždane kore čovjeka. Brodmannove areje (BA) 13-16 nedostaju, a predstavljaju brojeve koji vjerojatno označavaju inzulu (13 i 14 za kratke vijuge inzule, te 15 i 16 za dugu vijugu inzule). Polja 48-51 Brodmann nije opisao u čovjeka, već samo u lemura. Malo promijenjeno prema Brodmannu (1909) i Duvernoyu (1991). Pogled na lateralnu površinu lijeve hemisfere.
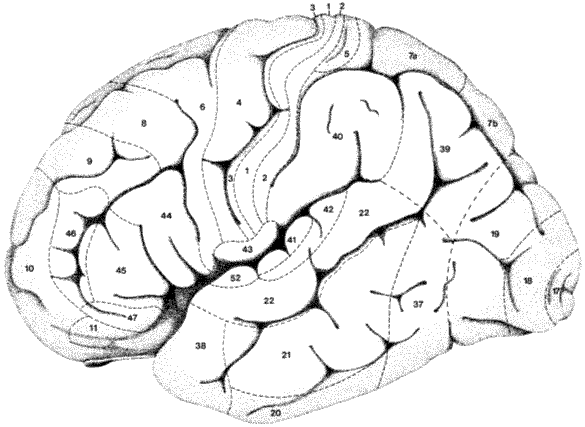
Slika 1b. Brodmannova citoarhitektonska mapa moždane kore čovjeka. Malo promijenjeno prema Brodmannu (1909) i Duvernoyu (1991). Pogled na donju i medijalnu površinu desne hemisfere.
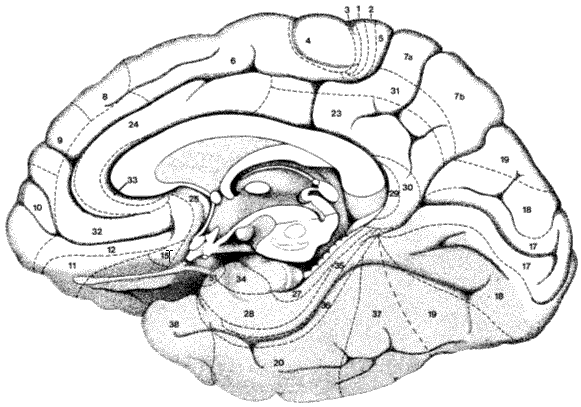
Nakon usporedne analize moždane
kore glavnih skupina sisavaca, Brodmann je uočio da između 6. i 8. mjesecatrudnoće
najveći dio (oko 90%) korteksa fetusa prolazi kroz stadij razvitka u kojem ima
6 slojeva (tzv. temeljni šestoslojni ontogenetski tip korteksa). Svaki dio odrasle
moždane kore koji se razvije iz tog temeljnog tipa naziva se isocortex ili neocortex
('nova', tj. evolucijski najmlađa moždana kora). Manji, preostali dio moždane
kore (oko 10%), nikad ne prođe kroz temeljni šestoslojni stadij, nastaje iz
drugačije razvojne osnove, pa se naziva allocortex. Alokorteks se dijeli na
dvoslojni paleokorteks, troslojni arhikorteks i petoslojni mezokorteks. U tijeku
kasnijeg razvitka, jedan dio izokorteksa zadrži šestoslojnu građu (homotipni
izokorteks), a manji se dio naknadno promijeni (heterotipni izokorteks), na
način da se broj slojeva smanji (tj. izgube se II i IV zrnati sloj pa nastane
agranularni korteks) ili poveća (II i IV sloj se naročito dobro razviju pa nastane
hipergranularni korteks ili koniokorteks). Homotipni korteks karakterističan
je za asocijacijska područja moždane kore (npr. BA9-11,
BA 5, 7, 39, 40,
BA18-22, 37). Agranularni korteks obuhvaća prednji dio inzule (BA13 i 14), precentralnu
vijugu (BA4, stražnji rub BA6) i prednji dio cingularne vijuge (BA24), a hipergranularnu
građu imaju primarno vidno (BA17), slušno (BA41 i 42) i somatosenzibilno (BA3,
1, 2) polje moždane kore.
Upotreba Brodmanovih areja može dovesti do nepreciznosti uslijed činjenice da
svi mozgovi nemaju ista neuroanatomska obilježja kao ona što ih je ilustrirao
Brodmann, te postojanja značajnih interindividualnih razlika u raspodjeli citoarhitektonskih
areja, čak i onda kada je morfologija vijuga i brazdi vrlo slična. Ostale strukturalne
mape moždane kore, kao npr. Campbellova koja sadrži 20 polja, von Economa i
Koskinasa koja ih ima 109, ili ona braće Vogt s preko 200 različitih citoarhitektonskih
dijelova, iako načelno u skladu s Brodmannovom podjelom, uslijed svoje premale
specifičnosti ili pak prevelike složenosti, nisu nikad doživjele širu upotrebu.
Za podjelu moždane kore prema funkcionalnim obilježjima vjerojatno su najzaslužniji
Broca, Campbell, Filimonoff, Yakovlev i Sanides, a ona je dovela do danas općeprihvaćene
podjele korteksa na 5 glavnih funkcionalnih tipova: 1) limbički, 2) paralimbički,
3) heteromodalni asocijacijski, 4) unimodalni asocijacijski, i 5) idiotipski
(primarni osjetni/motorički) korteks.
Limbički korteks se sastoji od kortikoidnih (neuroni organizirani u nakupine, ali se ne može jasno uočiti slojevita građa), paleokortikalnih i arhikortikalnih struktura. Kortikoidne strukture se nalaze u bazalnom telencefalonu. Iako se bazalni telencefalon često smatra subkortikalnom strukturom, neki njegovi dijelovi ipak se uključuju unutar granica moždane kore jer se nalaze na ventromedijalnoj površini moždanih hemisfera i tijesno su recipročno povezani s drugim limbičkim poljima. Stoga u bazalni telencefalon ubrajamo septalno područje, substanciju inominatu, amigdaloidni kompleks jezgara i prednju olfaktornu jezgru. U paleokorteks spadaju periamigdaloidni korteks, te prepiriformni i piriformni (ili primarni olfaktorni) korteks. Arhikorteks čini hipokampalna formacija što se sastoji od fascije dentate, četiriju polja Amonovog roga (CA1-CA4), prosubikuluma i subikuluma.
Slijedeći stupanj strukturalne
složenosti moždane kore uočava se u paralimbičkim područjima mozga koja se nazivaju
mezokorteksom. Ova područja nalaze se između alo- i izokorteksa i predstavljaju
postepeni prijelaz od prvog prema drugome. Paralimbička područja koja se nastavljaju
na alokorteks nazivaju se također i peri- ili jukstalokortikalna područja, dok
se ona što se naslanjaju na izokortikalna područja nazivaju pro- ili periizokortikalna
područja. Razgraničenja između ovih područja nikad nisu jasna i uključuju zone
postepenih prijelaza. U većini paralimbičkih područja, promjene na prijelazu
iz perialokorteksa u periizokorteks uključuju sve veći broj malih zrnatih neurona
najprije u sloju IV, a zatim i u sloju II, sublaminaciju i kolumnarizaciju sloja
III, razdvajanje sloja V od sloja VI i sloja VI od bijele tvari, te porast intrakortikalnog
mijelina, naročito uzduž sloja IV.
Paralimbički korteks sastoji se od 5 glavnih dijelova: orbitofrontalnog korteksa
(kaudalni dijelovi BA11 i 12, BA13), inzule (BA14-16), temporalnog pola (BA38),
parahipokampalnog korteksa koji uključuje presubikulum i parasubikulum (BA 27),
entorinalni (BA 28), pro- i peririnalni (transentorinalni) korteks (BA35 i 36),
te cingularnog kompleksa koji uključuje retrosplenijalni korteks, ventralni
cingulum i parolfaktorne areje (BA23-26 i BA29-33). Ovih pet paralimbičkih područja
čine neprekinuti (vanjski) prsten što okružuje medijalne i bazalne dijelove
moždanih hemisfera (tzv. unutrašnji prsten limbičkog režnja).
2.3. Unimodalni i heteromodalni asocijacijski korteks
Izokortikalna asocijacijska polja
dio su homotipnog korteksa, a dijele se na unimodalna i heteromodalna. Unimodalna
su vidna, slušna i somatosenzibilna asocijacijska polja. Neuroni koji ih čine
odgovaraju samo na osjetne podražaje jednog modaliteta, a glavne aferentne veze
dobivaju iz odgovarajućih primarnih osjetnih polja (ove veze se još nazivaju
i osjetnofugalne, okomite, unaprijedne ili 'nizvodne' veze) ili iz drugih unimodalnih
polja za istu vrst osjeta (osjetnopetalne, vodoravne, unatražne ili 'uzvodne'
veze). Unimodalno slušno polje zauzima gornji sljepoočni girus (BA22 i dijelovi
BA21), unimodalna somatosenzibilna polja zauzimaju gornji tjemeni režnjić (BA5
i 7) i prednje dijelove donjeg tjemenog režnjića (BA40), a unimodalna vidna
polja čine dijelovi zatiljnog režnja (BA18 i 19), fuziformnog i inferotemporalnog
korteksa (BA20 i 21), te spojnog područja zatiljnog, tjemenog i sljepoočnog
režnja (BA37). Unimodalna asocijacijska područja za njuh, okus i vestibularni
osjet nisu dobro poznata. Premotorička područja (prednji dio BA6 i BA8) zadovoljavaju
kriterije za unimodalna asocijacijska motorička polja, ali se obično ubrajaju
u primarna osjetna/motorička polja jer su građena od heterotipnog korteksa.
Heteromodalna komponenta asocijacijskog izokorteksa je definirana s tri glavna
obilježja: odgovori neurona nisu ograničeni na jedan modalitet, aferentne veze
primaju pretežno iz više unimodalnih, odnosno drugih heteromodalnih areja, a
deficit koji nastaje njezinim oštećenjem nikad nije ograničen na zadatke za
čije je izvršavanje dovoljan jedan modalitet. Stoga se ova polja nazivaju još
i poli-, supra-, multi- ili transmodalna, katkad još i 'zone prepokrivanja',
a nalaze se u prefrontalnom, naročito dorzolateralnom i prednjem orbitofrontalnom
korteksu (BA9-10, BA45-47, prednji dio BA11-12, prednji dio BA8), stražnjem
parijetalnom (stražnji dio BA7, BA39-40), i lateralnom sljepoočnom korteksu
(dijelovi BA37 i BA21), te dijelovima parahipokampalnog girusa (dijelovi BA36
i 37).
Iako se i unimodalna i heteromodalna polja ubrajaju u homotipični izokorteks,
između njih ipak postoje i neke relativno fine citoarhitektonske razlike. Općenito,
unimodalne areje su više diferencirane, naročito s obzirom na sublaminaciju
slojeva III i V, kolumnarizaciju sloja III, te granularizaciju slojeva IV i
II, što znači da je heteromodalni korteks sličniji paralimbičkom, pa predstavlja
stadij citoarhitektonske diferencijacije negdje između paralimbičkih i unimodalnih
područja.
Idiotipski korteks čine primarna
osjetna i motorička polja moždane kore. Ona su dijelovi heterotipnog korteksa,
a čine ih agranularno primarno motoričko polje precentralne vijuge (BA4 i stražnji
rub BA6), te hipergranularna primarna vidna (obje usne fisure kalkarine, BA17),
slušna (Heschlova vijuga, BA41 i 42) i somatosenzibilna (postcentralna vijuga,
BA3, 1 i 2) polja. Njušna, okusna i vestibularna polja nisu tako dobro izražena
u čovjeka i primata, pa su i odgovarajuće areje ovih osjeta manje, slabo diferencirane
i smještene bliže ili u sastavu limbičkih i paralimbičkih polja. Tako se primarni
okusni korteks nalazi u BA43, primarna vestibularna areja leži unutar stražnjeg
dijela Sylvijeve fisure na mjestu gdje se sljepoočni režanj spaja s inzulom
i parijetalnim režnjem, a primarni olfaktorni korteks u središtu limbičkog korteksa
na mjestu gdje konfluiraju inzularne, orbitofrontalne i temporopolarne areje.
Postoje dva suprotstavljena mišljenja o primarnim arejama. Jedno je mišljenje
da one predstavljaju elementarne (čak rudimentarne) dijelove moždane kore, a
drugo da su one zapravo vrlo napredne i najviše diferencirane areje, što podupire
i citoarhitektonska analiza. Naime, i primarni vidni i somatosenzibilni i slušni
korteks pokazuju najveći stupanj granularizacije i laminacije, dok primarni
motorički korteks pokazuje jedinstvenu 'makropiramidalnu' arhitekturu karakteriziranu
visokospecijaliziranim divovskim piramidnim neuronima (Betzove stanice).
3. Organizacija moždane kore i uloga transmodalnih areja u intermedijarnom procesiranju
Kao što je ukratko prikazano, korteks
moždanih hemisfera može se podijeliti u 5 temeljnih tipova koji pokazuju veliki
raspon citoarhitektonske diferencijacije. Kortikoidna, paleokortikalna i alokortikalna
područja koja čine središnje limbičke strukture opsežno su povezana dvosmjernim
vezama s hipotalamusom. Putem neuralnih i humoralnih (hormoni hipofize) mehanizama
hipotalamus nadzire cjelokupni endokrini i reproduktivni sustav, balans elektrolita,
razinu glukoze, bazalnu temperaturu, razinu metabolizma, tonus autonomnog živčanog
sustava, cirkadijane i druge bioritmove, imunološki status, te modulira iskustvo
i izražaj instinktivnih i nagonskih oblika ponašanja što su usmjereni na preživljavanje
i razmnožavanje (gladi, agresije, straha, bijega od opasnosti, žeđi, libida).
Ukorak s navedenim funkcijama hipotalamusa, limbički korteks ima ključnu ulogu
u regulaciji pamćenja, emocija i motivacije. Navedene funkcionalne specijalizacije
limbičkih struktura povezane su održanjem homeostaze organizma i s njom povezanih
djelovanja neophodnih za održanje jedinke i vrste.
Na strani 'suprotnoj' limbičkom korteksu nalaze se citoarhitektonski visoko
diferencirane i funkcionalno visoko specijalizirane primarne osjetne i motoričke
areje. Ovi dijelovi moždane kore najbliže su povezani s izvantjelesnim prostorom.
Primarni osjetni korteks omogućuje neophodni ulazak informacija iz okoline u
neuronske krugove moždane kore, dok primarni motorički korteks predstavlja zajednički
završni put svih motoričkih djelovanja koja omogućuju manipulaciju okoline i
promjenu pozicije unutar te okoline.
Iz primarnih osjetnih područja informacije najprije odlaze u modalitet-specifične
(unimodalne) asocijacijske areje. Svaka skupina unimodalnih areja prenosi zatim
'rezultate' svog modalitet-specifičnog procesiranja u limbički korteks (za pamćenje
i emocije), prefrontalni korteks (za radno pamćenje i izvršne funkcije), korteks
ispred i iza Sylvijeve brazde (za jezične funkcije), sljepoočni korteks (za
prepoznavanje objekata), stražnji parijetalni korteks (za prostornu pozornost)
i premotorički korteks (za osjetno vođenje pokreta i praksiju). Oštećenje unimodalnih
osjetnih asocijacijskih areja ili njihovih veza dovodi do dva glavna tipa poremećaja:
selektivnih percepcijskih deficita (kao što su npr. 'akromatopsija' i 'akinetopsija'),
čak i kad su druge funkcije u tom osjetnom modalitetu neoštećene, i 2) modalitet-specifičnih
'agnozija' i 'diskonekcijskih sindroma' kao što su npr. 'prozopagnozija' i 'čista
gluhoća za riječi' (vide infra).
Unimodalni, polimodalni i paralimbički korteks su interponirani između primarnih
osjetnih/motoričkih areja i limbičkog korteksa. Ove zone omogućuju povezivanje
osjetnih informacija, njihovo združivanje s strategijama djelovanja, te integraciju
iskustava s nagonima, emocijama i stanjima autonomnog živčanog sustava. Stoga
predstavljaju neuronske mostove koji posreduju između unutarnjeg i vanjskog
svijeta, tako da se potrebe organizma ostvaruju u skladu s mogućnostima i ograničenjima
koji vladaju u vanjskom svijetu. Pri tome su unimodalna i heteromodalna područja
najtješnje povezana s elaboracijom percepcije i motoričkog planiranja, dok paralimbički
korteks igra ključnu ulogu u kanaliziranju emocija i motivacije prema ponašanju
primjerenom mentalnom sadržaju i izvantjelesnim događajima.
Jedno od temeljnih obilježja moždane kore mozga čovjeka je postojanje većeg
broja sinapsi između podražaja i odgovora na podražaj, te između reprezentacije
internog milieura (na razini hipotalamusa) i reprezentacije vanjskog svijeta
(na razini idiotipskog korteksa). Ovi 'umetnuti' sinaptički slojevi predstavljaju
podlogu za tzv. intermedijarno (ili integrativno) procesiranje. Psihološki izraz
intermedijarnog procesiranja je kognicija, što uključuje različite izričaje
pamćenja, emocija, pozornosti, jezika, planiranja, promišljanja, uvida, itd..
Intermedijarno procesiranje 'štiti' kanale ulaznih osjetnih informacija i motoričkog
djelovanja od neposrednog motivacijom-vođenog utjecaja unutarnjeg milieura omogućujući
da identični podražaji dovedu do različitih odgovora u zavisnosti od konteksta
situacije, prošlog iskustva, trenutnih potreba i predmnijevanih posljedica.
Neuroni koji posreduju intermedijarno procesiranje nalaze se u unimodalnim,
heteromodalnim, paralimbičkim i limbičkim zonama moždane kore.
Sinaptička arhitektura intermedijarnog procesiranja oblikuje narav kognicije
i ponašanja. U životinjskih vrsta s jednostavnijim mozgovima, intermedijarno
procesiranje toliko je plitko da ne dopušta značajnije razlikovanje između pojavnosti
i važnosti. Automatska povezanost između podražaja i odgovora na podražaj utjelovljena
je u različitim oblicima instinktivnog ponašanja. Na primjer, puranica koja
je tek dobila potomstvo tretirat će svaki objekt koji se miče u blizini legla
kao neprijatelja, osim ako se ne glasa kao njeno mladunče. Ako joj se eksperimentalno
uništi osjet sluha, ona će bez iznimke ubiti sve svoje potomstvo. U ovakvim
oblicima ponašanja, fizičke osobine podražaja izazivaju unaprijed programirane
slijedove ponašanja relativno neosjetljive na kontekstualne razlike. Podloga
razvitka i uloga intermedijarnog procesiranja je transcendencija ovakvih direktnih
veza podražaj-odgovor, što omogućuje ponašanje vođeno kontekstualnom važnošću,
a ne samo fizičkim izgledom podražaja.
Unimodalne areje predstavljaju najpreciznije reprezentacije osjetnih iskustava,
sadrže inicijalne sinaptičke releje za intermedijarno procesiranje i izvanredno
su dobro razvijene u mozgu čovjeka. Nepostojanje direktnih veza koje bi povezivale
unimodalne areje jednog osjetnog modaliteta s drugima povećavaju sigurnost osjetnih
iskustava i usporavaju unakrsnu kontaminaciju sve dok se ne dovrši (kontekstualno)
prepoznavanje podražaja. Pored toga što kodiraju pojavna obilježja specifičnih
osjetnih doživljaja, ove areje mogu i npr. odrediti jesu li osjetna obilježja
složenih entiteta kao što su lica ili riječi identična, ili čak pohraniti ove
informacije u trajne engrame pamćenja. No, u odsutnosti pristupa informacijama
u drugim modalitetima, unimodalne areje nemaju mogućnost vođenja od riječi k
značenju, od fizionomije do prepoznavanja poznatih lica, ili pak od izoliranih
osjetnih događaja do cjelovitih iskustava. Za integraciju osjetnih iskustava
u kogniciju neophodna je participacija transmodalnih areja.
Obilježje koje definira transmodalnu areju je mogućnost podržavanja multimodalne
integracije, što znači odsutnost bilo kakve specifičnosti za bilo koji modalitet
osjetnog procesiranja. Prema ovoj definiciji svi su dijelovi heteromodalnog,
paralimbičkog i limbičkog korteksa također i transmodalni. Iako je važnost serijskog
procesiranja i multimodalne konvergencije od izuzetnog značenja u svim aspektima
mentalnog funkcioniranja, transmodalna integracija bila bi ipak vrlo ograničena
kada pored serijskog procesiranja od jedne prema drugoj sinaptičkog razini ne
bi također postojali i brojni paralelni putovi, unaprijedne i unatražne veze,
te mjesta divergencije i konvergencije. Glavna uloga transmodalnih areja je
zapravo ta da one predstavljaju 'direktorije' ('adresarije' ili u usporedbi
s računalom 'look-up' tablice) za povezivanje (a ne miješanje) modalitet-specifičnih
fragmenata informacija u cjelovita iskustva, sjećanja i misli. Transmodalne
areje uspostavljanju multimodalne asocijacije povezane s ciljnim događajem tako
da oblikuju sinaptičke veze koje pokazuju gdje su raspodijeljeni elementi s
tim događajem povezanih informacija. One dakle nisu 'centri' za pohranu konvergentnog
znanja već ključni 'čvorovi' za integraciju i pristup odgovarajućim raspodijeljenim
informacijama. To ujedno znači da predstavljaju i područja najveće vulnerabilnosti
za oštećenjima izazvane deficite u odgovarajućim domenama ponašanja i kognicije.
Iako je opisani način djelovanja svih transmodalnih areja sličan, svaka od njih
ima posebnu funkcionalnu specijalizaciju određenu sveukupnim obrascem njenih
neuronskih veza i fizioloških obilježja.
4. Glavne neuronske mreže moždane kore čovjeka
Primarni vidni korteks (BA17, V1)
zauzima gornju i donju usnu fisure kalkarine zatiljnog režnja. Zbog golim okom
vidljive bijele pruge (stria Gennari), što predstavlja debeli snop horizontalno
položenih mijeliniziranih aksona u sloju IVB, ovo je područje također poznato
pod imenom strijatalni korteks. Vidne informacije iz retine dolaze u strijatalni
korteks posredno putem projekcija neurona lateralnog koljenastog tijela (radiatio
optica). Cijelo vidno polje je mapirano na strijatalni korteks s velikom preciznošću.
Strijatalni korteks svake hemisfere prima vidne informacije iz vidnog polja
suprotne strane, pri čemu dorzalni dijelovi strijatalnog korteksa sadrže reprezentaciju
donjeg dijela vidnog polja, a ventralni gornjeg. Središnji (makularni, fovealni)
dio vidnog polja mapiran je na stražnjem dijelu strijatalnog korteksa i ima
najveću relativnu reprezentaciju, dok prednji dijelovi V1 sadrže reprezentacije
sve perifernijih dijelova vidnog polja. Osim za reprezentaciju vertikalnog meridijana,
strijatalni korteks u čovjeka nema kalozalne interhemisferičke veze (jednako
vrijedi i za sve druge primarne areje).
Oštećenje genikulostrijatalnih putova dovodi do karakterističnih deficita vidnog
polja unutar kojih je izgubljena svjesna percepcija objekata. Ipak, određena
rezidualna sposobnost posezanja za objektima u slijepom području vidnog polja
može biti sačuvana čak i kad bolesnik negira da je svjestan podražaja. Ovaj
fenomen rezidualnog vidnog kapaciteta ('blindsight') posljedica je intaktnosti
retinokolikularnih projekcija što se od donjih slojeva (S-I i S-II) lateralnog
koljenastog tijela projiciraju u gornje kolikule, a odavde u jezgre pulvinara
talamusa i na kraju asocijacijske vidne i parijetalne areje.
Vidne informacije se sastoje od mnogobrojnih fizičkih obilježja, npr. glavna
fizička obilježja boje su ton, jarkost i zasićenost. Iz ovih fizičkih obilježja
neuroni vidnog sustava 'ekstrahiraju' pojedine 'osjetne pokazatelje' ili vidne
submodalitete, npr. za boju su to luminancija i spektralni sastav svjetla reflektiranog
od objekata (npr. objekt koji reflektira najviše srednjih valnih duljina percipira
se kao zelen). Sama percepcija boja moguća je zahvaljujući cijelom nizu prilagodbi
i specijalizacija neurona vidnog sustava od kojih su najvažnije pigmenti čunjića
selektivno osjetljivih na različite valne duljine svjetlosti, te dvostruko-oponirajuća
receptivna polja V1 neurona. Naime, kad se gledaju objekti u različitim uvjetima
osvijetljenosti, kompozicija valnih duljina reflektiranog svjetla se mijenja,
a jedino je reflektancija konstantno svojstvo objekta. No, kad bi se percipirale
stalno različite boje u zavisnosti od uvjeta osvjetljenja, objekti se ne bi
mogli točno prepoznati i boja bi izgubila značaj biološkog signala. Neuroni
s dvostruko-oponentnim receptivnim poljima u V1 daju odgovor za konstantnost
gledanja boja. Naime, promjena uvjeta osvjetljenja na njih nema utjecaja budući
da jednako djeluje i na centar i na periferiju receptivnih polja, pa se proporcija
reflektancija pojedinih valnih duljina ne mijenja promjenom intenziteta ili
spektralne kompozicije incidentnog svjetla. To ujedno znači da se određena boja
može percipirati samo komparacijom reflektancija gledanog objekta i njegove
podloge (iz čega slijedi da se promjenom podloge može promijeniti i percepcija
boje objekta). Neuroni u V1 raspolažu samo pojedinačnim informacijama o razlikama
reflektancija, a svjesna se percepcija boje događa tek u V4 području složenom
usporedbom reflektancija dobivenih od svih triju sustava čunjića. Boja je, dakle,
kao i drugi vidni submodaliteti, vlasništvo mozga (što nastaje dugotrajnim svjesnim
procesom učenja, pa tako npr. iskusno 'oko' grafičara ili likovnog umjetnika
može naučiti razlikovati i do 250 nijansi boja), a ne fizičko obilježje objekta
(svjetlost je, sama po sebi, elektromagnetni val i nema 'boju').
Pored retinotopske lokacije, luminancije i reflektancije, V1 neuroni su selektivno
osjetljivi i na orijentaciju podražaja, smjer kretanja podražaja, binokularni
disparitet, duljinu i prostornu frekvenciju podražaja. Pri tome pojedini neuroni
mogu biti uključeni u obradu više različitih vidnih modaliteta, a obično se
tek povezivanjem više modaliteta 'procjenjuju' obilježja vanjskog svijeta. Da
bi izvršili veliki broj različitih vrsta ovakvih procjena, skupine neurona osjetljivih
na pojedine modalitete vidnih informacija organiziraju se u multidimenzionalne
mozaike kolumni, slojeva i citoarhitektonski diferenciranih 'metaboličkih' (citokrom-oksidaza
pozitivnih ili negativnih) 'otočića' vidnog korteksa. Iako još uvijek nije sasvim
jasno kako je moždana reprezentacija određenog opaženog obilježja (boje, oblika,
veličine, teksture, prozirnosti, krutosti/elastičnosti, 'zrcaljenja', uzajamnog
položaja, usmjerenosti, putanje itd.) povezana sa stvarnim fizičkim obilježjima
predmeta, znamo da su dobivena opažena obilježja specifične 'konstrukcije' ovakvih
skupina neurona, a svaka od njih utjelovljuje neku posebnu strategiju procesiranja
tako da u njima vidne informacije budu 'preuređene' na način da izravno služe
za prepoznavanje objekata i uočavanje njihovog položaja i kretanja u okolnom
prostoru. Ovakva funkcionalna specijalizacija nije ništa drugo nego ekonomizacija
i povećavanje učinkovitosti za izvršavanje navedenih operacija. Zbog brojnosti
fizičkih obilježja vidnih podražaja ne čudi da vidni korteks za njih upotrebljava
različite algoritme što onda uvjetuju posebnosti anatomskog ustroja i veza.
Zahvaljujući činjenici da se gotovo trećina moždane kore bavi obradom vidnih
informacija, možemo reći da moždane reprezentacije fizičkih obilježja vanjskog
svijeta uglavnom uspješno 'odslikavaju' i stvarna svojstva tog svijeta, iako
pod određenim uvjetima mogu od realnosti i znatno odstupati, pa nastaju vidne
iluzije.
Prijenos vidnih informacija od mrežnice do asocijacijskih područja moždane kore
sastoji se od dva velika usporedna sustava: P-sustava za opažanje boje i oblika
(koji započinje P ganglijskim stanicama retine čiji se aksoni projiciraju u
parvocelularne slojeve III-VI lateralnog koljenastog tijela) i M-sustava za
opažanje kretanja i akromatski vid (koji započinje M ganglijskim stanicama retine
čiji se aksoni projiciraju u magnocelularne slojeve I i II lateralnog koljenastog
tijela). Jedan sloj lateralnog koljenastog tijela prima vlakna iz samo jednog
oka: vlakna iz nazalne polovice retine suprotnog oka završavaju u I, IV i VI
sloju, a vlakna iz temporalne polovice istostranog oka završavaju u II, III
i V sloju. Razdvojenost P- i M-sustava očuvana je i na razini završetka genikulokortikalne
projekcije u IV sloju V1, jer se M-slojevi projiciraju u 4C podsloj, a P-slojevi
lateralnog koljenastog tijela u 4A i 4C podsloj. Pritom ova vlakna također čine
i izmjenične 'stupiće' (na okomitom presjeku), odnosno pruge (na tangencijalnom
presjeku) što odgovaraju aferentnom ulazu iz samo jednog oka ('okularno-dominantne
kolumne'). U poprečnom smjeru širine dvije okularno-dominantne kolumne nalazi
se pravilan slijed od oko 40-tak 'orijentacijskih kolumni' selektivno osjetljivih
na sve moguće orijentacije (0-360 , s razmakom od oko 9 između dviju susjednih
orijentacijskih kolumni) vidnog podražaja (tzv. 'hiperkolumna'). V1 se topografski
precizno projicira u V2 (BA18) i tek tu započinju brojne dvosmjerne veze P-
i M-sustava.
Od V1 i V2 područja započinju paralelni putovi koji se projiciraju u brojne
specijalizirane unimodalne vidne asocijacijske areje, kao što su V3, V4, V5
(MT), V6 (PO), V7 i V8, smještene većinom u peristrijatalnom korteksu, te dijelovima
lingvalnog, fuziformnog, donjeg i srednjeg temporalnog girusa (BA18-20, 37).
Daljnji okcipitofugalni tijek vidnih informacija poprima oblik divergentnih
multisinaptičkih putova od kojih su dva najizraženija: dorzalni (također poznat
pod imenom 'gdje' put) je usmjeren prema parijetalnom i frontalnom korteksu,
te ventralni ('što' put) usmjeren prema lateralnim temporalnim i temporopolarnim
vidnim asocijacijskim arejama i limbičkom korteksu. Pri tome je asocijacijsko
vidno polje V4 (vjerojatno i V8) u stražnjem dijelu ligvalnog i fuziformnog
girusa ključni posrednik u prijenosu informacija o boji i obliku iz P-puta u
asocijacijska polja sljepoočnog režnja, dok je V5 područje u dubini gornjeg
temporalnog sulkusa ključni posrednik u prijenosu vidnih informacija o kretanju
iz M-puta u parijetalni režanj.
Izolirano jednostrano oštećenje V4 areje dovodi do kontralateralnog gubitka
percepcije boja (hemiakromatopsije), bez poremećaja u vidnoj jasnoći, percepciji
kretanja ili prepoznavanju objekata. Oštećenja koja uzrokuju prekid izlaznih
veza V4 areje u korteks jezične mreže mogu dovesti do anomije za boje, tj. nesposobnosti
da bolesnik imenuje boje, iako je sama percepcija boja intaktna. Oštećenje V5
areje dovodi do hemiakinetopsije, tj. bolesnik ne može percipirati objekt koji
se kreće u kontralateralnoj polovici vidnog polja, iako su oštrina i percepcija
boja relativno sačuvane. Klinička disocijacija akromatopsije od akinetopsije
dokazuje da su projekcije primarnog vidnog korteksa organizirane i paralelno,
a ne samo serijski. Prisutnost paralelnog procesiranja povećava učinkovitost
procesiranja i složenost operacija za nekoliko redova veličine omogućujući istovremenu
obradu multiplih obilježja povezanih s vidnim događajima. Dodatnu potvrdu paralelnog
procesiranja daje i činjenica da svaka prestrijatalna vidna areja ima odvojeno
reprezentirano cijelo vidno polje.
U dubini sulkusa lunatusa i prednjem dijelu fuziformnog girusa (ispred V4) nalazi
se areja V3. Ova je areja uključena u konstrukciju oblika iz jednostavnijih
vidnih pokazatelja, a nastavlja se u susjedni lingvalni i donji okcipitalni
girus. Za razliku od V4 polja koja većinom prima informacije samo putem P-sustava
(a opsežno se projicira uglavnom u donji temporalni korteks) i V5 polja koje
prima informacije većinom samo putem M-sustava (a projicira se uglavnom u parijetalni
korteks), V3 područje prima vlakna i iz P- i iz M-sustava, a podjednako se projicira
i u temporalni i u parijetalni korteks. To je vjerojatno razlog zašto nikada
nije opisan potpuno izolirani gubitak percepcije oblika.
Obostrano oštećenje inferotemporalnih areja za prepoznavanje lica, areja za
prepoznavanje objekata, te jezičnih areja dovodi do specifičnih gubitaka sposobnosti
prepoznavanja lica ('asocijativna prozopagnozija'), složenih vidnih podražaja
(npr. mrtve prirode) (asocijativna vidna objektna agnozija) i vidnog prepoznavanja
riječi (čista aleksija). Oštećenja koja pak prekidaju veze između vidnih asocijacijskih
područja i limbičkih areja dovode do stanja vidne hipoemocionalnosti jer vidna
iskustva ne izazivaju prikladan emocionalni odgovor (npr. opisan je pacijent
koji je nakon oštećenja vidnoamigdaloidnog puta otkazao pretplatu na časopis
'Playboy' jer mu slike golih ljepotica nisu više izazivale erotske osjećaje)
i vidne amnezije (kada se ne mogu stvarati nova sjećanja u vidnom modalitetu).
Relativno jednostavniji aspekti percepcije lica (npr. sposobnost određivanja
da li dva lica imaju isti oblik) može u prozopagnozičara biti sačuvana, vjerojatno
zato što su 'niže' vidne asocijacijske areje ostale neoštećene. Pored lokalizacije
oštećenja u bolesnika sa sindromom prozopagnozije, i metode slikovnog prikaza
aktivacije mozga (fMRI) i elektrofiziološki evocirani odgovori u normalnih ispitanika
potvrđuju da su inferotemporalne areje specijalizirane za percepciju lica i
objekata smještene pretežno unutar prednjeg i srednjeg dijela fuziformnog girusa
(37 i 20), dok su areje specijalizirane za prepoznavanje riječi smještene
nešto lateralnije, tj. više okcipitotemporalno od fuziformne areje za lica (na
granici BA19 i BA37). S obzirom na kasnu pojavu pisanog jezika u filogenezi
čovjeka, areja za riječi gotovo sigurno nije genetski programirana. Vjerojatnija
je mogućnost da ona predstavlja modifikaciju podskupina neurona unutar skupina
specijaliziranih za prepoznavanje lica i objekata.
Neuroni inferotemporalnih areja za prepoznavanje lica jače se aktiviraju licima,
nego bilo kojim drugim objektima, a također su jače aktivirani intaktnim licima
u uspravnom položaju nego slikama namjerno 'zbrčkanih' lica u obrnutom položaju
(ali ne pokazuju razliku između poznatih od nepoznatih lica). Ove areje dakle
prepoznaju lica na kategoričkoj (generičkoj) razini, prije stadija prepoznavanja
o kojoj se eventualno poznatoj osobi radi, a informacije kojima raspolažu osiguravaju
izoliranu percepciju lica lišenu značenja ili konteksta. Sposobnost da ove modalitet-specifične
informacije aktiviraju relevantne multimodalne asocijacije koje dovode do prepoznavanja
poznatih lica zahtijeva posredovanje transmodalnih kortikalnih areja. U skladu
s navedenim, i fMRI istraživanja su potvrdila da ekspozicija nepoznatim licima
aktivira samo unimodalne vidne asocijacijske areje u području za lica fuziformnog
girusa, dok poznata lica također aktiviraju i transmodalne areje, naročito one
u srednjem sljepoočnom girusu. Transmodalne areje u srednjem sljepoočnom girusu
povezuju vidne reprezentacije lica s dodatnim asocijacijama (npr. imenom, glasom
i osobnim sjećanjima na tu osobu) što sve zajedno dovodi do prepoznavanja. Iako
bolesnici s asocijativnom prozopagnozijom ne mogu gledanjem prepoznati poznata
lica, prepoznavanje postaje moguće kad dobiju informaciju u ne-vidnom modalitetu,
npr. kad čuju glas dotične osobe. Naime, ova auditorna informacija dovodi do
transmodalne aktivacije kroz unimodalni slušni korteks, što pak dovodi do aktivacije
drugih raspodijeljenih asocijacija koje dovode do prepoznavanja. Nadalje, i
lice koje nije svjesno prepoznato može katkada izazvati fiziološki emocionalni
odgovor ako je oštećenje dovelo samo do prekida veza između vidnih asocijacijskih
areja i srednjeg sljepoočnog girusa, a ne i između vidnih asocijacijskih i limbičkih
areja.
Dok prozopagnozija predstavlja nemogućnost prepoznavanja jedinstvenih primjeraka
vidno kodiranih kategorija objekata (za što je potrebno puno više informacija
u odnosu na određivanje generičkih obilježja objekta), asocijativna vidna objektna
agnozija predstavlja poremećaj koji je smješten 'niže' od kategoričkog prepoznavanja.
Bolesnici s ovim sindromom ne mogu ni imenovati poznati objekt, niti opisati
njegovu prirodu i upotrebu. Dok prozopagnozičar može reći da je lice lice, a
olovka olovka, bolesnik s objektnom agnozijom to nije u stanju (ali može imati
sačuvanu sposobnost određivanja jesu li su dva objekta identična). Druga razlika
između prozopagnozije i objektne agnozije temelji se na sustavima pamćenja koji
ih posreduju. Naime, iako se oba poremećaja češće vide nakon obostranih oštećenja,
katkad nastaju i nakon jednostranog oštećenja. U tom slučaju je prozopagnozija
obično rezultat oštećenja u desnoj, a objektna agnozija u lijevoj hemisferi.
Ova disocijacija nastaje jer desna hemisfera ima veću ulogu u aktivaciji autobiografskih
sjećanja, što ukazuje da je generičko prepoznavanje poznatih objekata više dio
semantičkog znanja, dok je prepoznavanje poznatih lica i objekata više povezano
s vlastitim iskustvom.
Asocijativne agnozije postoje i u auditornom i taktilnom modalitetu. Bolesnici
s auditornom objektnom agnozijom ne mogu povezati zvonjenje telefona ili sirene
ambulantnih kola s odgovarajućim objektom koji je taj zvuk proizveo, iako su
neke elementarne slušne sposobnosti obično sačuvane. Ovaj sindrom odraz je prekida
veza između unimodalnih slušnih areja specijaliziranih za kodiranje auditornih
svojstava poznatih objekata i transmodalnih areja koje koordiniraju njihovo
multimodalno prepoznavanje. Oštećenja koja dovode do auditorne agnozije tipično
zahvaćaju auditorni asocijacijski korteks, obično desne hemisfere. Slušni analog
prozopagnozije je fonagnozija. Ona odražava nesposobnost prepoznavanja poznatih
glasova, potencijalno i u odsutnosti drugih slušnih deficita. Ovaj rijetki sindrom
može biti posljedica diskonekcije areje specijalizirane za identifikaciju boje
glasa u unimodalnom asocijacijskom korteksu i heteromodalnog lateralnog sljepoočnog
korteksa.
Taktilna agnozija je nesposobnost prepoznavanja objekata palpacijom (u odsutnosti
bilo kakvog drugog somatosenzibilnog ili afazičkog deficita). Teorijski se može
razlikovati od astereognozije (vide infra) tako da se pokaže da bolesnik s taktilnom
agnozijom (za razliku od astereognostičara) može nacrtati objekte koje palpira,
čak i kad palpacija ne dovede do prepoznavanja. Iz ovog se može zaključiti da
je astereognozija aperceptivni deficit, dok je taktilna agnozija asocijativni
deficit prepoznavanja objekta putem somatosenzibilnog sustava. Odgovorno oštećenje
obično je smješteno u inzuli ili stražnjem parijetalnom korteksu, tako da prekida
projekcije iz somatosenzornog asocijacijskog korteksa u transmodalne areje lateralnog
temporalnog korteksa.
Navedene modalitet-specifične agnozije naglašavaju važnost osjetnofugalnih putova
u procesima prepoznavanja i daju neuroanatomski temelj razlikovanju percepcije
od prepoznavanja.
Vidna integracija je stupnjevit proces, a svaki njen stupanj eksplicitno pridonosi
percepciji ili prepoznavanju. Integracija vidne percepcije proizlazi iz povratnih
veza i vremenskog sinhroniciteta izbijanja neurona (sinhronicitet sam po sebi
zahtijeva povratne veze). Budući da povratne veze nisu modularne, već difuzne,
i sam proces vidne integracije je nemoguće lokalizirati. Budući da se vidni
korteks sastoji iz mnogobrojnih areja, diskretna oštećenja ne interferiraju
toliko sa samim procesom integracije, koliko s oštećenjem pojedinih vidnih submodaliteta.
Opsežna povezanost vidnih areja također omogućuje 'pretvaranje' jednog submodaliteta
iz drugoga (npr. oblik iz pokreta ili oblik iz sjene i dubine).
Vidne iluzije zorno ilustriraju da je vidna percepcija aktivan i kreativan proces
u kojem neuroni vidnog sustava transformiraju vidne podražaje u mentalne složaje
koristeći se pritom i brojnim nesvjesnim pretpostavkama i očekivanjima koje
dijelom potječu iz iskustva, a dijelom iz samog ustroja pojedinih areja uključenih
u obradu vidnih informacija. Tako se npr. u V2 može snimiti aktivnost neurona
koji reagiraju na virtualne (nepostojeće) linije, a ovi neuroni mogu biti i
selektivno oštećeni. Navedenu činjenicu dodatno potvrđuju snovi i halucinacije,
jer predstavljaju vidne percepcije koje nastaju i u odsutnosti vidnih podražaja.
Iako različite osjetilne mape moždane
kore imaju različitu metriku (npr. vidna mapa je retinotopna, somatosenzibilna
somatotopna, zvučna cefalotopna, a kinestetska gravitotopna), njihove se 'koordinate'
preračunavaju u zajednički koordinatni sustav čije je središte točka smještena
na sredini između očiju (egocentar). Koordinatna mapa personalnog (onaj dio
prostora što ga zauzima tijelo), peripersonalnog (onaj dio prostora što izravno
okružuje tijelo i unutar je dosega ruke) i ekstrapersonalnog (sav preostali
dio prostora o kojem se osjetilne obavijesti dobivaju uglavnom na temelju vida,
sluha i njuha) prostora nalazi se u moždanoj kori stražnjeg parijetalnog korteksa,
u koju konfluiraju vlakna vidnih, slušnih, somatosenzibilnih i vestibularnih
areja. Stražnji parijetalni korteks sadrži također i heteromodalne areje koje
posreduju multimodalnu integraciju, a topografski se dijeli u: 1) gornji parijetalni
režnjić, što sadrži somatosenzibilni asocijacijski korteks (BA5 i prednji dio
BA7), te heteromodalni korteks (stražnji dio BA7), 2) donji parijetalni režnjić,
što sadrži supramarginalni (BA39) i angularni (BA40) girus, a ima obilježja
heteromodalnog korteksa osim prednjeg ruba supramarginalnog girusa koji pripada
somatosenzibilnom asocijacijskom korteksu, 3) duboki intraparijetalni sulkus,
što ima osjetno-motorička obilježja u prednjem, a vidno-prostorna u stražnjem
dijelu, 4) medijalni parijetalni korteks, što u prednjem dijelu sadrži somatosenzibilni
asocijacijski (BA5 i 7), a u stražnjem heteromodalni korteks (BA7 i 31). Pored
toga što stražnji parijetalni režanj ima bogate veze s osjetnim asocijacijskim
područjima, on je opsežno povezan i s čeonim područjem za očne pokrete, premotoričkim
korteksom, gornjim kolikulima, parahipokampalnim girusom, te paralimbičkim arejama
uključujući orbitofrontalni korteks, inzulu i naročito girus cinguli.
Za veliku neuronsku mrežu što čini sustav prostorne pozornosti (orijentacije)
najvažnija su tri međusobno povezana epicentra: stražnji parijetalni korteks
iznad i ispod parijetalnog sulkusa (BA 5 i 7), čeono područje za očne pokrete
(BA8) i girus cinguli (BA24). Parijetalna komponenta specijalizirana je za stvaranje
privremene reprezentacije lokacija u prostoru i njihovu transformaciju u ciljeve
usmjeravanja pozornosti na suprotnoj polovici ekstrapersonalnog prostora. Čeona
polja za očne pokrete upravljaju istražujućim i orijentirajućim usmjeravanjem
pogleda prema zanimljivom objektu (jednostrano oštećenje dovodi do konjugirane
devijacije očiju prema strani oštećenja), dok girus cinguli raspodjeljuje pozornost
s obzirom na motivaciju (prije usmjeravanja pogleda ili posezanja rukom, treba
odlučiti je li objekt uopće vrijedan pozornosti). Dodatne ključne komponente
ove goleme mreže nalaze se u talamusu (naročito intralaminarnim jezgrama), strijatumu
i gornjim kolikulima.
Balint je 1909. godine opisao pacijenta J.K. s tri karakteristična poremećaja:
optičkom ataksijom (nemogućnosti zagledavanja u određenu točku, odnosno pokazivanja
određenog smjera ili predmeta), paralizom pogleda (nesposobnošću usmjeravanja
pozornosti na više od jednog objekta), te sustavnim otklonom polja pozornosti
prema desnoj strani, a obdukcijom je utvrđena obostrana ozljeda stražnjeg parijetalnog
režnja. Stoga je zaključeno da tjemeni režanj sadrži mapu ekstrapersonalnog
prostora što služi za 'aktivno gledanje', tj. usmjeravanje pogleda, pokreta
i pozornosti na temelju vidnih informacija. Dok nakon opsežnih obostranih oštećenja
tjemenog režnja nastaje Balintov sindrom, u slučaju jednostranih i manjih oštećenja
nastaju samo neke njegove komponente. Tako sindrom jednostranog zanemarivanja
('hemi-neglect') nastaje gotovo isključivo nakon oštećenja desnog tjemenog režnja
(zbog dominantnosti desne hemisfere za prostornu pozornost, oštećenje lijevog
tjemenog režnja rijetko dovodi do sličnih simptoma). Primjerice, bolesnik ne
češlja lijevu stranu glave, ne brije lijevu stranu lica, ne jede hranu iz lijeve
polovice tanjura, ne piše po lijevoj polovici papira, ne odijeva lijevu ruku
i nogu (apraksija odijevanja), ne može prepoznati predmet pod neobičnim kutom
gledanja (konstrukcijska apraksija), itd.. Teži oblici ovakvog oštećenja mogu
dovesti i do astatognozije (nemogućnosti određivanja položaja dijelova tijela
u prostoru) i asomatognozije (bolesnik tvrdi da lijeva strana nije njegova,
pa npr. hoda samo s desnom nogom, a lijevu samo pasivno vuče za njom; opisan
je čak i slučaj bolesnika koji je stalno ispadao iz kreveta jer je pokušavao
iz njega istjerati 'tuđu' nogu). Nešto blaži oblik oštećenja desnog tjemenog
režnja naziva se sindrom jednostrane nepozornosti ('hemi-inattention'), a najblaži
oblik se opisuje kao simultagnozija (tek kad se dva osjetna podražaja jave istodobno
na obje strane vidnog polja, bolesnik ne obraća pozornost na podražaj što se
javi na strani suprotnoj od oštećenja).
Prepoznavanje lica i objekata u bolesnika s Balintovim sindromom obično je normalno,
ali katkad može također biti oštećeno, a naziva se 'aperceptivna prozopagnozija'
jer odražava deficit prostorne integracije samog vidnog podražaja. Suprotno
asocijativnoj prozopagnoziji, bolesnik s aperceptivnom prozopagnozijom obično
je sposoban odrediti jesu li su npr. dva lica potpuno identična.
Gerstmann je 1924. godine opisao bolesnika s: 1) desnostranim gubitkom taktilne
diskriminacije i astereognozijom (neprepoznavanjem) prstiju i drugih objekata
dodirom, 2) nerazlikovanjem lijevog i desnog (lijevo-desna dezorijentacija),
te 3) nemogućnošću izražavanja misli pisanjem - disgrafijom (ali uz sačuvanu
mogućnost kopiranja napisanih riječi), disleksijom (poremećajem čitanja) i diskalkulijom
(poremećajem računanja), a obdukcija je pokazala izdvojenu ozljedu lijeve angularne
vijuge (BA39). Pored činjenice da je Gerstmannov sindrom ukazao na ulogu tjemenog
režnja u prepoznavanju predmeta opipavanjem, on je također prvi skrenuo pozornost
na spoznajne funkcije lijevog tjemenog režnja (čitanje, pisanje, računanje),
što istovremeno participiraju u velikoj neuronskoj mreži jezičnih sposobnosti.
4.3. Mreža jezičnih sposobnosti s epicentrima u Wernicke i Broca području
Jezik omogućuje obradu i prijenos
misli i iskustava pomoću arbitrarnih simbola - riječi. Wernickeovo i Broca područje
čine dva epicentra mreže jezičnih sposobnosti. Zbog lateralizacije jezičnih
funkcija, u većine se ljudi deficiti ove mreže vide samo nakon oštećenja lijeve
hemisfere. Dodatne komponente mreže nalaze se u asocijativnim područjima čeonih,
sljepoočnih i parijetalnih režnjeva, strijatumu i talamusu.
Wernickeovo područje obuhvaća stražnju trećinu gornjeg sljepoočnog girusa (BA22),
dijelove susjednih heteromodalnih područja (BA39 i 40) i dijelove srednjeg sljepoočnog
girusa (BA21), a specijalizirano je za leksičke i semantičke aspekte jezika.
Oštećenje Wernickeovog područja dovodi do dvosmjernog neurološkog deficita:
bolesnik ne može imenovati objekt koji mu se pokaže, ni pokazati objekt kad
mu se kaže njegov naziv. Ovaj nalaz je posljedica činjenice da je Wernickeovo
područje transmodalna areja koja koordinira recipročne interakcije između osjetnih
reprezentacija riječi i simboličkih asocijacija koje im daju značenje. Njeno
oštećenje stoga ne dokida reprezentacije riječi samih, već čini nemogućim njihovo
dekodiranje u bilo koji osjetni modalitet, i obrnuto, onemogućuje kodiranje
(povezivanje) osjetnih percepata u odgovarajuće oblike riječi. Analogija organizacije
jezične mreže s mrežom za prepoznavanje lica i objekata očigledna je, a slično
je i objašnjenje ekvivalenata njihovih oštećenja.
Dva su tipa jezičnih asocijativnih agnozija. Čista aleksija (sljepoća za riječi)
nastaje kad su areje koje kodiraju vidne oblike riječi diskonektirane od vidnih
areja ili ne mogu komunicirati s Wernickeovom arejom. Ovo se obično događa nakon
lezija koje istovremeno oštećuju V1 areju lijeve hemisfere i splenijum (stražnji
dio korpusa kalozuma). Oštećenje splenijuma ne dopušta transfer vidnih informacija
iz neoštećene desne hemisfere u areje za kodiranje vidnih oblika riječi, a ni
u transmodalne areje za jezik lijeve hemisfere (tako ove areje ostaju diskonektirane
i od ipsi- i od kontralateralnog vidnog ulaza). No, bolesnik s čistom aleksijom
nije slijep budući da informacije o objektima i licima iz lijeve polovice vidnog
polja prepoznaje desnim heteromodalnim sljepoočnim korteksom. On također nije
ni afazičan budući da Wernickeova i druge jezične areje primaju informacije
u slušnom modalitetu. No, kada mu se kaže da čita, djeluje kao da je potpuno
nepismen jer transmodalne jezične areje ne dobivaju informacije o riječima u
vidnom modalitetu.
U slučaju čiste gluhoće za riječi unimodalni slušni korteks je lišen ulaza auditornih
informacija ili ne može komunicirati s odgovarajućim transmodalnim arejama.
Čista gluhoća za riječi može se vidjeti nakon obostranog ili unilateralnog oštećenja
sljepoočnog režnja za jezik dominantne hemisfere (obično lijeve) što prekida
ipsilateralne i transkalozalne slušne veze iz ostalih komponenata jezične mreže.
Kako su im obično dijelovi primarnog slušnog korteksa i slušnih asocijacijskih
areja intaktni, ovakvi bolesnici nisu gluhi i mogu bez poteškoća interpretirati
većinu zvukova iz okoline. Oni također nisu ni afazični, niti pokazuju poteškoće
u čitanju, pisanju i govoru budući su im jezične areje većinom neoštećene. No,
kad čuju govor, reagiraju kao da se radi o nekom tuđem jeziku jer ga uslijed
činjenice da im slušne informacije ne dolaze do Wernickeove areje ne mogu razumjeti.
Somatosenzibilni ekvivalent navedenih poremećaja je čista agrafestezija (ili
somatosenzibilna aleksija). Ovakvi bolesnici ne mogu u taktilnom modalitetu
prepoznati slova (iako im je normalno imenovanje objekta u ruci i somatosenzibilno
prepoznavanje jesu li dva slova što ih drže u ruci identična). Odgovorno oštećenje
zahvaća stražnji parijetalni korteks lijeve hemisfere i prekida veze između
asocijacijskog somatosenzibilnog korteksa i Wernickeove areje.
Area Broca je čeono središte jezične mreže. Ona uključuje premotorički korteks
u BA44 i susjedne heteromodalne čeone areje BA45-47. Areja Broca specijalizirana
je za artikulatorne, sintaktičke i gramatičke aspekte jezika. Proučavanje bolesnika
s žarišnim oštećenjem ove areje ukazuje da ona ima ključnu ulogu u prevođenju
neuralnih oblika riječi u njihove artikulatorne slijedove, ali također i u slaganju
riječi u govor koji ima značenjski prikladnu sintaktičku strukturu. Ovakva uloga
u slaganju riječi u skladu je s drugim specijalizacijama premotoričkog korteksa.
Areja Broca pokazuje aktivaciju (fMRI) u zadacima koji u neurološki normalnih
ispitanika traže dešifriranje značenja sintaktički složenih rečenica, te zadacima
koji zahtijevaju transformacije grafema u foneme, čak i kad se artikulatorno
djelovanje samo zamišlja.
Oštećenja koja diskonektiraju BA6 od stražnjih multimodalnih komponenti jezične
mreže (naročito onih u supramarginalnom girusu, BA40) uzrokuju ideomotornu apraksiju
(nemogućnost izvođenja složenih motoričkih zadataka, npr. salutiranja, odnosno
nesposobnost da se nakon verbalnog upita pantomimom objasni upotreba objekta),
iako bolesnik ne mora imati poteškoća u izvođenju istih pokreta nakon pokazivanja
pokreta (u prvom slučaju) ili kad mu se pokaže stvarni objekt (u drugom slučaju).
Postoje dvije vrste 'modalitet-specifičnih' apraksija. U vidnoj apraksiji bolesnici
ne mogu pantomimom objasniti upotrebu objekta koji im je prikazan vizualno (a
na verbalni upit mogu). Kod taktilne (palpatorne) apraksije bolesnici ne mogu
objasniti upotrebu objekta kad ga drže u ruci (u odsutnosti vidne kontrole).
Ovi deficiti nastaju zbog oštećenja koja prekidaju vidne, odnosno somatosenzibilne
projekcije u premotorička i suplementarna motorička područja.
Izraz afazija koristi se za opisivanje poremećaja procesiranja jezika koji su
uzrokovani disfunkcijom pojedinih dijelova mozga. Afazija nastaje uslijed prekida
procesa dvosmjerne komunikacije koja uspostavlja korespodenciju između misli
i jezika. Afazični bolesnici nisu u stanju pretvarati neverbalne slike koje
čine misli u jezične simbole i gramatičke relacije koje čine jezik, a u većini
slučajeva, i obrnuti smjer procesa je defektan: kad bolesnik čuje riječ ne može
od nje sačiniti neverbalnu sliku koja odgovara značenju te riječi. Po analogiji
s drugim mrežama, afazija je poremećaj razumijevanja, a ne percepcije (npr.
gluhoća oštećuje razumijevanje jezika kroz modalitet sluha, ali ne i kroz vidni
i taktilni modalitet). Afazija također nije motorički poremećaj (to je npr.
disartrija, koja remeti artikulaciju, ali ne kompromitira uobličavanje jezika),
ni rezultat poremećenog procesa mišljenja (kao npr. schizophrenia, kod koje
je sam lingvistički aparat sačuvan). Afazija također nije vezana isključivo
za jezike koji se temelje na govoru, jer i gluhonijeme osobe koje koriste znakovni
(vidnomotorički) jezik mogu postati afazične (kao rezultat fokalnog oštećenja
u lijevoj hemisferi dominantnoj za znakovni jezik), ili pak jezike koji su utemeljeni
na pismu (pa afazija može nastati i u osoba koji se koriste ideogramskim pismom).
Afazija narušava različite aspekte jezične obrade: 1) morfosintaksu (gramatiku),
2) semantiku (značenje riječi), i 3) fonologiju (izgovor glasova) i 4) pragmatiku
(uporabu jezika). Često su nekoliko ili svi aspekti jezika kompromitirani u
istog pacijenta, a različite kombinacije daju i različite sindrome afazije.
Najčešći uzroci afazija su ozljeda glave, moždani udar, Alzheimerova bolest
i tumori.
Broca afazija (motorička, ekspresivna, netečna ili 'prednja' afazija). U 'klasičnoj'
Broca afaziji oštećenje je opsežno, a zahvaća area Broca (BA44 i 45), tj. pars
triangularis i pars opercularis lijevog donjeg čeonog girusa, priležeće dijelove
lijevog čeonog (BA9, 47, 46), lijevog parijetalnog (BA40, 39, 7) i somatosenzibilnog
korteksa (donji dijelovi BA3, 1, 2 i 4), kao i pripadajuću bijelu tvar i bazalne
ganglije (glavu kaudatusa i putamen). Većina ih ima i desnu hemiparezu, obično
jače izraženu u desnoj ruci nego nozi. U Broca afaziji su oštećeni neuronski
krugovi odgovorni za spajanje fonema u riječi i spajanje riječi u rečenice (vremenski
slijed lingvističkih komponenti), te gramatičku strukturu rečenica i pravilnu
upotrebu gramatičkih morfema. Stoga je govor bolesnika s Broca afazijom težak
i spor, pauze između riječi su dugačke, nema melodične modulacije koja karakterizira
normalan govor, jednom riječju govor je netečan ('nonfluent'). Bolesnici s klasičnom
Brokinom afazijom razumiju značenje rečenica, ali ih ne mogu ponoviti, čemu
se često i sami čude (a budući da su svjesni deficita, postaju depresivni).
Agramatizam je jedno od najizraženijih obilježja, pa nastaje telegrafski govor,
npr. 'Ići...ja...kući...sutra'. No, iako govore malo, pacijenti s Brokinom afazijom
prenose smislene ideje. Razumijevanje pročitanog im je poremećeno uglavnom zbog
nemogućnosti razumijevanja gramatički važnih riječi (veznika, prepozicija, članova,
itd.). Ovi bolesnici također često imaju poremećen izgovor fonema, npr. miješaju
/b/ za /p/, iako im je njihovo prepoznavanje neoštećeno. Oštećenje nedominantne
(desne) hemisfere u području korespodentnom Broca areji dovodi do ekspresivne
disprozodije, tj. nemogućnosti artikulacije naglaska, intonacije i melodije
(ritma) govora.
Potpuno ograničen defekt središnjeg dijela Broca areje ili samo vlakana točno
ispod nje dovodi do afemije, što je zapravo poremećaj motoričkog izvršavanja
govora, a ne pravi jezični deficit. Afemija zato nije prava afazija jer se ovi
pacijenti mogu normalno izraziti pisanim jezikom, te u potpunosti razumiju i
pisani i govorni jezik. Iako im je govor slabo artikuliran, izbor riječi i sintaksa
su normalni.
Wernickeova afazija (osjetna, receptivna, tečna ili 'stražnja' afazija). Wernikeova
afazija nastaje zbog oštećenja stražnjeg dijela lijevog slušnog asocijacijskog
korteksa (BA22), a često su zahvaćeni i dijelovi BA37, 39 i 40. Wernickeova
afazija dovodi ne samo do nemogućnosti razumijevanja jezika u svim modalitetima,
nego i do nemogućnosti izražavanja misli značenjski prikladnim riječima. Bolesnici
s Wernickeovom afazijom podsjećaju na klasičnu Broca afaziju samo u smislu nemogućnosti
ponavljanja rečenica, slaganja fonema i imenovanja, a u svim drugim aspektima
su različiti jer im je: 1) govor tečan ('fluent'), tj. bolesnik govori bez napora,
melodično, normalnom ili češće povećanom brzinom, 2) zbog čestih pogrešaka u
izboru fonema i riječi sadržaj izgovorenog im je često nerazumljiv, 3) imaju
velike teškoće u razumijevanju rečenica koje čuju, 4) rijetko imaju bilo kakav
motorički poremećaj. Pogrešne riječi koje bolesnici s Wernickeovom afazijom
izgovore umjesto onih koje su inicijalno zapravo 'htjeli' često su slične po
značenju, npr. kažu 'poglavica' umjesto 'predsjednik' (ovaj poremećaj naziva
se verbalna ili semantička parafazija). Iz istog razloga oni često rabe riječi
općeg značenja, npr. 'stvar', u zamjenu za one do kojih ne mogu doći. Bolesnici
s Wernickeovom afazijom nemaju problema s produkcijom individualnih fonema,
no često pomiču slijed glasova ili glasovnih nizova, tako da neke dodaju ili
izbace što poremeti fonemski izgled željene riječi, npr. 'pimarida' umjesto
'piramida' (fonemska parafazija). Kad se fonemske parafazije javljaju učestalo,
nastaju neologizmi (neologijske parafazije), npr. 'hipidomateus' za 'hipopotamus'.
Iako puno govore, ovi bolesnici često ne prenose nikakvu ideju ili smisao. Neki
bolesnici s Wernickeovom afazijom su više 'gluhi za riječi', a neki više 'slijepi
za riječi'. Razumijevanje izgovorenog im je obično nešto bolje nego pročitanog.
Oštećenje nedominantne (desne) hemisfere u području korespodentnom Wernickeovoj
areji dovodi do osjetne disprozodije, tj. nemogućnosti percepcije naglaska,
intonacije i melodije (ritma) govora.
Kondukcijska afazija. Bolesnici s kondukcijskom afazijom mogu razumjeti i izgovoriti
jednostavne rečenice. No, oni ne mogu ponoviti rečenice doslovce, jer nisu u
stanju učinkovito slagati foneme (nastaje puno fonemičkih parafazija). Stoga
je oštećeno čitanje naglas, no čitanje radi razumijevanja je relativno dobro
sačuvano (uslijed činjenice da im je temporalno područje za razumijevanje jezika
uglavnom neoštećeno). Ova vrst afazije nastaje zbog oštećenja lijevog supramarginalnog
girusa (BA40) ili lijevog primarnog slušnog korteksa (BA41 i 42) i inzule. U
oba slučaja Wernickeova je areja sačuvana, a oštećen je snop vlakana koji povezuje
audiovizualna asocijacijska područja (BA22, 39 i 40) s area Broca za govor (BA44
i 45). Ovaj snop (fasciculus arcuatus) prolazi ispod angularnog i supramarginalnog
girusa i frontoparijetalnog operkuluma, a služi za povezivanje fonema u morfeme
i riječi što je neophodna operacija za vokalizaciju riječi, bez obzira da li
se one percipiraju u auditornom modalitetu ili generiraju interno.
Globalna afazija. Bolesnici s globalnom afazijom imaju gotovo potpuni gubitak
sposobnosti razumijevanja jezika i produkcije govora, dakle kombinirana obilježja
Wernickeove i Broca afazije. Slobodni ('propozicijski') govor je obično reduciran
na nekoliko riječi ili rečenica. Česta je upotreba stereotipnih riječi u pokušaju
da se iznese ideja ili misao.
Transkortikalne afazije. Ovi se poremećaji razlikuju od svih drugih afazija
po tome što je ponavljanje rečenica normalno. Motorički oblik transkortikalne
afazije nastaje uslijed oštećenja lijevog čeonog režnja ispred ili iznad area
Broca, a osjetni oblik zbog oštećenja korteksa na parijetotemporalnom spoju
(angularni girus, srednji i gornji temporalni girus) podsjeća na Wernickeovu
afaziju, osim što bolesnici mogu ponoviti što im se kaže. Miješana transkortikalna
afazija (katkad se naziva i izolacija areje za govor), vrlo je rijetka, a može
se smatrati analogom globalne afazije, s tom razlikom da ovakav bolesnik može
ponoviti ono što mu se kaže. No, nakon ponavljanja ('eha'), nema daljnje 'konverzacije'.
Akinetski mutizam je potpuni prestanak bilo kakvog verbalnog izričaja. Može
nastati zbog obostranog oštećenja čeonog režnja (suplementarne motoričke areje,
Broca areje), mezencefalona, infarkta lijevog talamusa. Mutizam može i ne mora
biti povezan s afazijom (npr. može ga uzrokovati teški laringitis), no u prisutnosti
mutizma ne može se utvrditi afazija.
Anomička afazija je najčešća od svih afazija, a u tipičnim primjerima, ovi bolesnici
su nesposobni pronaći pravu riječ, bilo u spontanom govoru ili u pisanju. Kad
nastupi akutno, najčešće je oštećen temporoparijetalni spoj, kad je blaga teško
se lokalizira.
Subkortikalna afazija nastaje zbog infarkta ili krvarenja u kaudatus, putamen,
talamus, suplementarnu motoričku areju i druge dijelove teritorija koji opskrbljuje
prednja cerebralna arterija.
Aferentna vlakna dolaze u hipokampus
najvećim dijelom indirektno preko entorinalnih područja. Entorinalna moždana
kora glavno je mjesto konvergencije putova iz svih važnijih asocijativnih i
osjetnih područja neokorteksa, a informacije iz tih područja dolaze u entorinalni
korteks također posredno iz transmodalnih areja. Lateralni dio entorinalnog
korteksa (28b) čini glavno stjecište vlakana iz 'najviših' vidnih, a u manjoj
mjeri i slušnih i somatozenzoričkih asocijativnih područja. Stoga se čini da
entorinalni korteks šalje hipokampalnoj formaciji 'sažetak zbivanja', tj. svega
onoga što se događa u moždanoj kori. Medijalni dio entorinalnog korteksa (BA28a)
više je pod utjecajem kortikoidnih, paleo- i arhikortikalnih limbičkih struktura
(koje koje odražavaju unutrašnje stanje organizma). Sveukupno, anatomsku povezanost
hipokampusa s neokorteksom, kakva je potvrđena i snimanjem sinaptičkih potencijala,
može se opisati na slijedeći način: neokorteks -> parahipokampalni girus
-> površni (II i III) slojevi entorinalnog korteksa -> zrnate stanice
fascije dentate -> CA3 područje hipokampusa -> CA1 područje hipokampusa
-> subikulum -> duboki (V i VI) slojevi entorinalnog korteksa -> neokorteks.
Eferentna vlakna entorinalnog korteksa odlaze iz njegovih dubokih slojeva natrag
u prepiriformnu (BA51), prelimbičku (BA32), infralimbičku (BA25), agranularnu
inzularnu i cingularnu (BA23 i 24), retrosplenijalnu (BA29), peririnalnu (BA35
i 36), stražnju orbitofrontalnu (BA13) i sljepoočnu (BA20, 22, 38 i 41) moždanu
koru. Peririnalna moždana kora se npr. dalje projicira u prefrontalnu (BA9 i
46), sljepoočnu (BA20-22), tjemenu (BA7) i zatiljnu (BA18) moždanu koru. Ove
projekcije navedene su kao ilustracija činjenice da se rezultati hipokampalnog
procesiranja diseminiraju po brojnim kortikalnim područjima, uključujući i primarna
osjetna područja (njušno - BA51, slušno - BA41), osjetno-specifična asocijativna
područja (vidno - BA19-21, slušno - BA 22, somatosenzoričko - BA7), kao i transmodalna
asocijativna područja (BA9, 13, 23, 38 i 46). Sažeto se stoga može reći da je
hipokampus povezan s glavnim transmodalnim asocijativnim područjima moždane
kore s jedne, te limbičkim i subkortikalnim strukturama s druge strane. Dodatne
kritične komponente ove mreže nalaze se u paralimbičkim arejama, hipotalamusu,
limbičkom dijelu talamusa i limbičkom strijatumu.
Ključni pomak u razumijevanju uloge hipokampusa u procesima pamćenja donio je
slučaj 27 godina starog pacijenta H.M., kojemu je u cilju otklanjanja neukrotive
epilepsije Scoville 1953. godine odstranio obostrano dvije trećine prednjeg
dijela hipokampusa, veći dio parahipokampalnog girusa, prednji dio sljepoočne
moždane kore, unkus i amigdala. Nakon operacije, koja je konačno dokinula epileptičke
napadaje, H.M. se nije mogao sjetiti prostorija u kojima je živio, imena svojih
prijatelja, trenutnog datuma niti godine, hrane koju je pojeo prilikom zadnjeg
obroka, pa čak niti puta do nužnika. Istovremeno, nije došlo do nikakvog oštećenja
drugih intelektualnih funkcija niti osobina ličnosti (kvocijent inteligencije
bio mu je i dalje normalan: prije operacije 1953. godine iznosio je 104, a poslije
operacije, 1955. godine 112, 1962. godine 117, 1981. godine 100). H.M. je naime
izgubio sposobnost zadržavanja novih činjenica i događaja (anterogradna amnezija),
iako je Milner ustanovila da se još uvijek mogao prisjetiti činjenica i događaja
koji su se dogodili prije operacije (retrogradno pamćenje), i to onih davnih
bolje i lakše (s vremenom čak i jasnije) nego onih što su se dogodili neposredno
prije operacije. Nasuprot tome, tzv. kratkotrajno pamćenje (traje oko 40-60
sekundi) bilo mu je u okviru normalnih granica. Tako je H. M. bez problema rješavao
male vidne i taktilne labirinte za čije rješenje nije trebalo više od minute,
a također je dobro rješavao križaljke. Praktično jedini gubitak bila je potpuna
nesposobnost pretvorbe kratkotrajnog u dugotrajno pamćenje. H.M. je tako svakodnevno
razgovarao sa osobljem bolnice, ali se nakon toga nije mogao sjetiti ni s kim,
niti što je razgovarao. Samo u nekoliko rijetkih prilika mogao se prisjetiti
emotivno jako nabijenih činjenica, npr. potvrdio je da se donekle sjeća očeve
smrti koja se dogodila prije dva mjeseca od upita, te se kroz još kraće razdoblje
mogao prisjetiti smrti pape Ivana XXIII i atentata na tadašnjeg predsjednika
Kennedyja. Ovaj tzv. Claparedeov fenomen boljeg pamćenja emocionalno nabijenijih
činjenica uočava se i u drugih amnestičnih pacijenata, uključujući i one s Korsakovljevim
sindromom.
Da je za značajan gubitak pamćenja - globalnu amneziju, neophodno obostrano
odstranjenje hipokampusa, uvidjelo se kad je u samo dvoje (bolesnici P.B. i
F.C.) od 90 slučajeva jednostrane sljepoočne lobektomije zabilježeno teže oštećenje
pamćenja slično onome H.M.-a. U prilog ovoj tvrdnji govori i činjenica da su
u jednom (P.B.) od ta dva slučaja nakon smrti obdukcijom ustanovljene degenerativne
promjene suprotnog hipokampusa (izgled ovih promjena ukazivao je da su najvjerojatnije
nastale zbog hernijacije hipokampusa kroz otvor tentorija još u vrijeme porođaja).
Ovaj izvorni nalaz o nužnosti obostranog oštećenja za izazivanje trajne globalne
amnezije kasnije su potvrdili i drugi autori (zbog čega su se operacije s obostranim
odstanjivanjem sljepoočnih režnjeva oko 1968. godine prestale izvoditi), a s
vremenom su se pojavili i drugi bolesnici čija je klinička slika manje-više
sličila onoj H.M.-a (npr. pacijent Boswell koji je obolio od herpes simplex
encefalitisa, a opisali su ga Damasio i Damasio, te pacijent R.B. koji je preživio
ishemički napadaj uslijed komplikacija pri operaciji premošćivanja koronarne
arterije, a kojeg je pratio i pet godina nakon tog događaja obducirao Zola-Morgan).
Ova je činjenica također postala putokaz neurokirurzima da prije svake hipokampektomije
pomoću amobarbitala (paralizira sljepoočni režanj one strane na kojoj je kroz
karotidnu arteriju injiciran) i jednostavnih testova zapamćivanja ustanove očuvanost
sljepoočnog režnja suprotne strane (tzv. Wada test). No, i u slučaju jednostrane
hipokampektomije može se psihološkim testovima dokazati blaže oštećenje pamćenja
koje nije globalno, već sadržajno određeno. U slučaju odstranjenja lijevog hipokampusa
mogu se otkriti poteškoće u zapamćivanju verbalnih sadržaja, dok u slučaju odstranjenja
desnog hipokampusa dotične osobe imaju otežano zapamćivanje vidnih sadržaja,
npr. teško procjenju jesu li već bile na određenom mjestu ('deja vu'), te imaju
blaže poteškoće u zapamćivanju orijentacije, prostornih odnosa i lica.
Za razliku od anterogradne amnezije koja se nastavila nesmanjeno i 49 godine
nakon operacije (tj. do danas) i retrogradne amnezije za činjenice i događaje
koji su se dogodili nekoliko mjeseci do nekoliko godina prije operacije, bolesnik
H.M. mogao je usvajati motoričke i perceptivne sposobnosti (zrcalno pisanje,
rješavanje labirinta i sl.), što je ukazivalo da postoje različite vrste pamćenja
koje nisu jednako oštećene. Tako je postulirano da je posebna vrsta pamćenja
ono koje se odnosi na svjesno prisjećanje činjenica i događaja, i koje ovisi
od integriteta hipokampusa, dok je drugi oblik plastičnosti živčanog sustava
onaj koji obuhvaća određene sposobnosti koje ne uključuju neuronske krugove
koji posreduju svjesno doživljavanje i ne ovise od entorinalno-hipokampalnih
struktura. Budući da se prvonavedeni oblik pamćenja može izgovoriti, odnosno
opisati, ono je nazvano deklarativno (još i eksplicitno ili faktičko) pamćenje
('knowing that memory'), dok je drugonavedeni oblik, kod kojeg iskustvo mijenja
ponašanje na nesvjesnoj razini, dobio naziv nedeklarativno (još i implicitno,
proceduralno ili refleksivno) pamćenje ('knowing how memory'). Deklarativno
pamćenje obično se sastoji od asocijacija istovremenih podražaja, a dijeli se
na semantičko i epizodičko. Semantičko (još i značenjsko ili referentno) pamćenje
obuhvaća pamćenje općeprihvaćenih činjenica i koncepata (npr. 'formula vode
je H2O', ili 'glavni grad Hrvatske je Zagreb') za koje nije potrebno kontekstualno
prisjećanje (npr. kad smo se prvi put susreli s vodom ili kad smo prvi put čuli
za Zagreb), pa ga se još naziva i mentalnim leksikonom. Neovisno je od događaja
u životu osobe. Testovi kojima se doznaje o stanju semantičkog pamćenja obuhvaćaju
značenje riječi, mogućnost imenovanja uobičajenih objekata ili dobro poznatih
geografskih, povjesnih ili npr. aritmetičkih činjenica. Epizodičko (ili autobiografsko)
pamćenje predstavlja informacije o autobiografskim događajima, kao i vremenske
i prostorne asocijacije za navedene događaje, a nastaje već kao rezultat samo
jedanput doživljenog događaja. Izjava 'Jučer sam u Hrvatskom institutu za istraživanje
mozga vidio preparate hipokampusa' primjer je epizodičkog pamćenja, ali činjenica
da se institut za istraživanje mozga nalazi na Šalati spada u semantičko pamćenje.
Oštećenje hipokampusa značajno oštećuje epizodičko pamćenje tj. mogućnost zapamćivanja
novih informacija, dok su različiti aspekti semantičkog pamćenja tipično sačuvani.
Semantičko pamćenje očigledno je u mozgu višestruko reprezentirano tako da je
svaka takva reprezentacija povezana s modalitetom putem kojeg je to znanje dobiveno.
Ova činjenica temelji se na analizi ponašanja bolesnika koji imaju žarišna oštećenja
sljepoočnog režnja, a pokazuju selektivno oštećenje za navođenje obilježja npr.
životinja i biljaka, kad bi im se rekla njihova imena. Nasuprot tome, kad bi
im se prikazale slike tih istih životinja i biljaka, oni odmah znaju navesti
njihova obilježja. Zbog ovakve višestruke reprezentiranosti semantičko je pamćenje
manje zahvaćeno oštećenjem različitih dijelova limbičkog sustava. Semantičko
pamćenje je izgleda organizirano kao složena mreža međusobno povezanih koncepata,
i to tako da su koncepti koji dijele puno istih zajedničkih obilježja bolje
povezani od onih koji dijele manje zajedničkih obilježja. Npr. pas i lav se
oba kategoriziraju kao životinje jer dijele zajedničke obilježja kao što su
biti živ, biti pokretan, moći se razmnožavati, ali se mogu i razlikovati međusobno
jedan od drugog npr. po obilježjima pripitomljenosti, veličine i oblika. Koncepti
i asocijacije koje čine semantičko znanje nalaze se transmodalnim arejama moždane
kore.
Nasuprot deklarativnom, nedeklarativno pamćenje je 'automatsko', jer ne ovisi
o svijesti, pažnji, ni spoznajnim funkcijama tj. stupnju znanja, a u amnestičnih
bolesnika je obično sačuvano. Iskazuje se poboljšanim izvođenjem određenih zadataka
i polako se kumulira tijekom brojnih pokušaja (npr. hodanje, zakapčanje gumba,
učenje gramatike), a temelji se na plastičnosti samih osjetnih i motoričkih
sustava (kao posljedica toga, može se izvrsno proučavati i na različitim modelima,
bilo u kralježnjaka ili dapače beskralježnjaka). Kod viših vrsta bitno je uočiti
da se brojnim ponavljanjima deklarativnog sadržaja pamćenja ono pretvara u nedeklarativno
pamćenje, što je posredovano kortikostrijatalnim putovima. Npr. učenje upravljanja
automobilom najprije uključuje lingvistički proces svjesnog deklariranja (izgovaranja
naglas ili 'u sebi') radnji koje je potrebno izvršiti, a ponavljanjem ono prelazi
u nesvjesnu motoričku aktivnost. U nedeklarativno pamćenje spadaju motoričke,
percepcijske i kognitivne vještine i navike, razina adaptacije, 'priming' (sposobnost
dovršenja prethodno prikazane slike ili riječi; zanimljivo je da bolesnici s
globalnom, limbičkom amnezijom, imaju ovu sposobnost sačuvanu, usprkos negiranja
da su ikad prije vidjeli testirane slike ili riječi), te asocijativni (koji
nastaju kao odgovor na spregu dvaju ili više događaja, a dijele se na jednostavno
ili klasično i operantno kondicioniranje) i neasocijativni (koji nastaju iz
iskustva sa samo jednom vrstom događaja, a dijele se na habituaciju, senzitizaciju
i 'imprinting') oblici učenja.
Nalaz neoštećenog kratkotrajnog pamćenja u bolesnika H.M. posljedica je činjenice
da je morfološki supstrat kratkotrajnog pamćenja u asocijativnim područjima
neokorteksa, pa je onda moguća i tzv. obrnuta disocijacija, tj. slabo kratkotrajno
uz očuvano dugotrajno pamćenje. Prvi slučaj koji je potvrdio navedenu pretpostavku
opisali su Warrington i Shallice 1969. godine, a kasnije su opisani i drugi
slični slučajevi. Radilo se o bolesniku K.F. koji nakon pada s motocikla nije
mogao zapamtiti niti dva broja (a kod ponavljanog testiranja jedva i jedan)
ili dva slova (normalni opseg kratkotrajnog pamćenja je oko 6-7 riječi ili brojeva),
iako je njegovo dugotrajno pamćenje za svakodnevne događaje bilo normalno (npr.
mogao se bez poteškoća prisjetiti 9 od 10 asocijacija nakon razdoblja od 6 sati).
Koliko je vremena potrebno funkcionalno sposobnom (neoštećenom) hipokampusu
za izvršenje konsolidacije, tj. pretvorbe kratkotrajnog u dugotrajno pamćenje?
Drugim riječima, koliko je razdoblje nakon stjecanja iskustva, poslije kojega
oštećenje hipokampusa ne bi oštetilo prisjećanje? U eksperimentima koje su prije
petnaestak godina izvršili Zola-Morgan i Squire približili smo se sasvim blizu
odgovoru na ovo pitanje. Majmuni su trenirani odgovarati na zadatak povezivanja
objekata, tako da su im bili predstavljeni slučajni parovi različitih objekata,
ali je samo jedan od objekata bio povezan s nagradom u obliku hrane. Objekti
su bili u skupinama od po 20, a skupine su bile pokazivane u različitim razmacima
s obzirom na vrijeme izvršenja operacije: na razmaku od 16, 12, 8, 4 i 2 tjedna
prije obostranog odstranjenja hipokampusa, girusa dentatusa i entorinalnog korteksa.
Rezultat je bio slijedeći: objekte koje su majmuni naučili razlikovati 2 do
4 tjedna prije lezije, prepoznavali su samo slučajno. Dakle, u ovim slučajevima
hipokampalne strukture nisu bile dovoljno dugo intaktne nakon pokazivanja podražaja
da bi bile zapamćene. Nasuprot tome, oni majmuni kojima je načinjena navedena
operacija 8 i 12 tjedana nakon naučenog raspoznavanja izvodili su jednako točno
zadatak kao i kontrole. To znači da je hipokampusu bilo potrebno 6-8 tjedana
da bi pohranio, tj. konsolidirao naučeni odgovor u druge dijelove korteksa i
tako ga učinio neovisnim od hipokampalnih struktura. Početno se kodiranje informacija
koje stižu iz multimodalnih asocijativnih područja putem parahipokampalnih aferentnih
vlakana događa unutar hipokampalnih krugova za vrijeme kad se snime theta sinaptički
potencijali (već u svom originalnom članku iz 1954. godine kad su otkrili ritmičku
sporu aktivnost, tj. theta ritam, Green i Arduini opisali su da oštećenje septalnih
jezgara dokida hipokampalni theta ritam), dok se konsolidacija nakon toga događa
u neuronskim krugovima unutar i izvan hipokampusa za vrijeme oštrih valova.
Za vrijeme eksplorativne aktivnosti (theta valovi) putem neurona površnih slojeva
entorinalnog korteksa informacije se prenose u iz neokorteksa u hipokampus,
dok se za vrijeme konzumatornog ponašanja i sporovalnog spavanja (oštri valovi)
sinhroniziranim izbijanjem 'autoasocijatinog matriksa' Schafferovog sustava
povratnih kolateralnih veza neurona CA3 područja (jedan CA3 neuron može davati
i po desetak kolaterala!), te CA1 i subikularnog područja, signali preko dubokih
slojeva entorinalnog korteksa prenose natrag u neokorteks (tj. hipokampalna
autoasocijativna mreža se 'prazni' u neokorteks u kratkom razdoblju - tzv. 'hipokampalna
kompresija vremena').
Pod amnestičnim pacijentom obično se podrazumijeva čovjek s oštećenim anterogradnim
pamćenjem, jer je isključivo retrogradna amnezija izuzetno rijedak fenomen.
Tako u više od 1000 dobro dokumentiranih slučajeva traume glave nije pronađen
niti jedan slučaj retrogradne amnezije kojeg nije pratila i anterogradna amnezija.
Zapravo postoji samo jedan dobro opisan i dokumentiran slučaj isključivo retrogradne
amnezije, kod kojeg je došlo do oporavka anterogradnog, ali ne i retrogradnog
pamćenja. Ovaj 36-godišnji čovjek imao je 1981. godine otvorenu frakturu lubanje
desnog parijetookcipitalnog i sljepoočnog područja uz kompresiju lijevog mezencefalona
u području tentorijalnog otvora. Nakon operacije najznačajniji nalaz na CT snimci
bio je uski tračak smanjene gustoće tkiva koji se protezao od ventralnog tegmentalnog
područja do pontomezencefaličke granice, a uključivao je i fasciculus medialis
telencephali (medial forebrain bundle). Nakon dvije godine od operacije došlo
je do potpunog oporavka anterogradnog, ali ne i retrogradnog pamćenja (što je
inače uobičajen slučaj u amnestičnih pacijenata), jer je on i dalje tvrdio da
je star između 16 i 18 godina i nije se mogao sjetiti niti da je oženjen, niti
da ima djecu, niti prijašnjih zaposlenja. Ova vrst poremećaja pamćenja nazvana
je mezencefalička amnezija.
Osim limbičke i mezencefaličke amnezije, i oštećenja nekih drugih struktura
mozga mogu dovesti do amnezije. Najčešća je tzv. diencefalička amnezija: oštećene
strukture su mamilarna tijela i mediodorzalna (ponekad i prednja) jezgra talamusa.
Uzrok poremećaja obično je Korsakovljev sindrom, stanje koje najčešće nastaje
uslijed kroničnog alkoholizma i posljedičnog manjka vitamina skupine B, no može
to biti i poremećaj cirkulacije, koloidna cista trećeg ventrikla ili tumor.
Poremećaji pamćenja u ovim slučajevima nastaju zbog oštećenih procesa pažnje,
nerazumijevanja značenja riječi i drugih procesa koji se odvijaju u tijeku učenja,
a ne zbog nemogućnosti prisjećanja. Zato bolesnici s diencefaličkom amnezijom
uče vrlo sporo, ali ono što nauče teško zaboravljaju (za razliku od bolesnika
s oštećenjem hipokampusa koji zapravo uče brzo, ali brzo i zaboravljaju). I
kod diencefaličke amnezije je anterogradna amnezija puno jača nego retrogradna,
što potvrđuje da se mehanizmi za prizivanje u svijest davnih događaja razlikuju
od mehanizama koji posreduju svježije događaje. I ovi pacijenti imaju normalan
'priming'.
Do amnezije također može dovesti i oštećenje bazalnog telencefalona (najčešći
uzrok degeneracije neurona bazalnog telencefalona je Alzheimerova bolest, no
to može biti i npr. ruptura aneurizme prednje komunikantne arterije). Jedino
oštećenje medijalne septalne jezgre dovodi do direktnog i jasnog oštećenja pamćenja
(jer medijalna septalna jezgra aktivira hipokampus), dok oštećenja drugih dijelova
bazalnog telencefalona samo posredno i slabije oštećuju ovaj proces.
Entorinalna moždana kora i hipokampus su među najranije zahvaćenim strukturama
mozga patoanatomskim promjenama za vrijeme Alzheimerove bolesti, a obrazac deficita
pamćenja sličan je onome nakon hipokampektomije, tj. onome u bolesnika s globalnom
amnezijom. Razlog tome je činjenica da neuropatološke promjene (neurofibrilarna
degeneracija i odlaganje beta-amiloida) dovode do prekida veza između entorinalno-hipokampalnih
struktura s neokortikalnim područjima (diskonekcijski sindrom). Zato je slabljenje
zapamćivanja najraniji, najizraženiji i najstalniji znak Alzheimerove bolesti.
U prvom, ranom stadiju karakterističan je gubitak epizodičkog pamćenja, naročito
usvajanja novih sadržaja, što je praćeno i progresivnom deterioracijom prisjećanja
već usvojenih epizodičkih sadržaja. Poteškoće u pronalaženju prave riječi (anomička
disfazija) u tijeku spontanog govora obično su prvi karakteristični znak uslijed
čega pacijent zaobilaznim putem nastoji reći što je naumio (pa nastaje logoreja).
U drugom stadiju, sposobnosti učenja, prepoznavanja najbliže okolice i rodbine,
te semantičko pamćenje postaju sve jače zahvaćene (uslijed difuznih oštećenja
brojnih, prvenstveno asocijativnih neokortikalnih područja s pohranjenim semantičkim
sadržajima), dok pamćenje davnih događaja ne mora biti u potpunosti izgubljeno
(za utvrđivanje poremećaja semantičkog poremećaja najkorisnijim se pokazao test
u kojem se pacijentu kaže da nacrta sat: tipične konceptualne greške u crtanju
sata u osoba oboljelih od Alzheimerove bolesti uključuju sat nacrtan bez brojeva
ili su brojevi krivo raspoređeni, neispravno crtanje kazaljki ili pisanje stvarnog
vremena brojkama umjesto crtanja kazaljki i slično, što sve ukazuje da su obilježja
koja karakteriziraju sat i značenje samog sata kao pokazivača vremena u njih
izgubljeni). Još uvijek je dobro sačuvano proceduralno pamćenje tj. pacijent
zadržava sposobnost učenja novih motoričkih obrazaca. U posljednjem, trećem
stadiju, deklarativno pamćenje je u potpunosti izgubljeno (i epizodičko i semantičko),
a također se počinje gubiti i proceduralno.
Uloga amigdaloidnog kompleksa jezgara, u novije je vrijeme ponovno ispitana
selektivnim odstranjivanjem u majmuna. Iako je jedno kraće vrijeme (do konca
80-tih) vladalo mišljenje da i amigdala igraju podjednako važnu ulogu u pamćenju
kao i hipokampus, 1989. godine je Zola-Morgan sa suradnicima potvrdio izvorni
nalaz Narabayashija iz 1963. godine, odnosno da potpuno i obostrano odstranjenje
amigdala ne dovodi u majmuna do nikakvog značajnijeg oštećenja pamćenja. Inicijalno
je do ove 'zabune' vjerojatno došlo jer su teža oštećenja pamćenja u eksperimentalnih
životinja kojima su odstranjena i amigdala i hipokampus, u odnosu na one kojima
je odstranjen samo hipokampus, bila rezultat oštećenja korteksa (što je pri
takvim operacijama teško izbjeći), a ne amigdala. Dodatna potvrda dobivena je
snimanjem metaboličke aktivnosti mozga pomoću radioaktivno obilježene 2-deoksi-D-glukoze
u majmuna. U nizu vidnoprostornih zadataka odgođenog odgovora vidjela se utilizacija
glukoze, između ostalog, i u području hipokampalne formacije, ali ne i amigdala.
Za razliku od hipokampalne formacije koja prima aferentna vlakna iz hetero-
i transmodalnih dijelova moždane kore, amigdala imaju veze samo sa uni- i heteromodalnim
dijelovima asocijativnog korteksa, tj. primaju manje 'prerađene' osjetne informacije.
Najvažnije uloge amigdaloidnog kompleksa jezgara mogu se sažeti u asociranje
između različitih modaliteta i asociranje između podražaja i nagrade (samo pri
ponovljenim susretima s podražajem).
Istovremeno oštećenje i hipokampusa i amigdala naziva se Klüver-Bucy sindrom,
prema autorima koji su ga 1937. godine prvi opisali kod majmuna. Sindrom se
u čovjeka, pored izražene amnezije, sastoji od gubitka osjećaja straha (u 100%
slučajeva), oralnog ispitivanja objekata (u 98% slučajeva), hiperseksualnosti
(79%), obraćanja pažnje na svaki vidni podražaj (hipermetamorfoze) (77%), gubitka
svake emocionalnosti (74%), vidne agnozije i hipoemocionalnosti (42%) i bulimije
(u 28% slučajeva). U novije je vrijeme pomoću modernih metoda slikovnog prikaza
aktivacije mozga utvrđeno da oštećenje naročito lijevog amigdaloidnog kompleksa
jezgara remeti prepoznavanje i prosudbu intenziteta straha iskazanog izrazom
lica (iako sposobnost prepoznavanja dotičnog lica ostaje sačuvana).
Čeoni režnjevi mozga čovjeka doživjeli
su tijekom filogeneze zadivljujuće povećanje, te zauzimaju otprilike trećinu
moždane kore. Korteks čeonog režnja može se podijeliti na 3 funkcionalna područja:
1) 'Motoričko-premotoričko' koje uključuje BA4 i 6, suplementarnu motoričku
areju (medijalni dio BA6), čeona polja za očne pokrete (lateralni dio BA6),
te dijelove area Broca (BA44). U zavisnosti od točne lokalizacije, oštećenje
ovog područja dovodi do slabosti mišića i promjene mišićnog tonusa (sve do fleksijskih
kontraktura), oslobađanja primitivnih refleksa (hvatanja, sisanja, itd.), inkontinencije,
akinezije, mutizma, aprozodije, apraksije, Broca afazije i nekih motoričkih
komponenti sindroma jednostranog zanemarivanja; 2) 'Paralimbičko' područje smješteno
je u ventralnim i medijalnim dijelovima čeonog režnja i sadrži korteks prednjeg
cinguluma (BA23 i 32), ravnog girusa (BA25) i stražnjeg orbitofrontalnog područja
(BA11-13); 3) 'Heteromodalno' područje je najopsežnije, a obuhvaća BA9 i 10,
prednje dijelove BA11 i 12, te BA45-47. Ovo područje ima dvosmjerne veze sa
svim drugim heteromodalnim, te gotovo svim unimodalnim, paralimbičkim i limbičkim
arejama moždane kore. Kao i u slučaju parijetotemporalnog heteromodalnog korteksa,
i ovdje su neki neuroni selektivno osjetljivi na jedan ili samo neke modalitete,
a 'pomiješani' su s onima najbrojnijima što imaju multimodalna obilježja. Izrazi
'prefrontalni korteks' i 'sindrom čeonog režnja' zapravo se odnose samo na paralimbičke
i heteromodalne dijelove čeonog režnja.
Vjerojatno prvi dobro dokumentirani opis selektivnog oštećenja čeonog režnja
u čovjeka bio je slučaj Pineas Gagea kojega je 1868. godine opisao Harlow. Gage
je bio pouzdan predradnik u izgradnji bostonske željeznice. Prilikom utiskivanja
baruta u zemlju pomoću željezne šipke, zbog prejake kompresije baruta, došlo
je do eksplozije zbog koje mu je katapultirana željezna šipka prošla kroz čeone
režnjeve mozga. Po preživljavanju ovog incidenta, Gage je iskazivao dramatične
promjene u razmišljanju i osobnosti, iako su mu osjetno-motoričke funkcije ostale
relativno sačuvane. Inspiriran Golzovim eksperimentima kirurške ablacije neokorteksa
psa iz 1890. godine, Burkhardt je 1892. godine odstranio različite dijelove
korteksa u 6 schizofrenih bolesnika s halucinacijama, no zbog kriticizma medicinskih
autoriteta njegovog vremena se do 1935. godine 'psihokirurške' operacije uglavnom
više nisu izvodile. Te su godine Jakobsen i Fulton objavili da su nakon oštećivanja
prefrontalnog i frontalnog korteksa (lobotomije) uspjeli agresivne čimpanze
učiniti mirnima (štoviše, u njih više nisu mogli izazvati eksperimentalnu neurozu),
a Penfield je opisao učinke desne prefrontalne lobektomije koju je načinio svojoj
sestri. Nakon akutnog postoperativnog razdoblja, on je uočio da su njene sposobnosti
planiranja i brzine razmišljanja postale vrlo ograničene, iako su joj socijalne
vještine i druge kognitivne sposobnosti ostale relativno sačuvane. Pa ipak,
i pored ovih 'nuspojava', neki su liječnici i kirurzi bili oduševljeni navedenim
postupkom. Misleći da će time ukloniti paranoju i opsesivno-kompulzivni poremećaj
('rekurentne obrasce misli') u svojih bolesnika, Moniz i Lima razvili su kiruršku
proceduru 'leukotomiju' koja je, nakon trepanacije lubanje na nekoliko mjesta,
obuhvaćala bilateralno presijecanje vlakana koja povezuju prefrontalni korteks
s talamusom (pedunculus frontalis thalami). Iako je paraplegični Moniz (nakon
što ga je u leđnu moždinu nastrijelio bivši pacijent), nedugo zatim otišao u
mirovinu, ovaj je postupak doživio veliku popularizaciju zahvaljujući Freemanu
i Wattsu (tzv. 'Freeman-Wattsova standardna procedura'). Kad je Watts razočaran
u učinke operacije prekinuo suradnju s Freemanom, 1945. godine je Freeman razvio
tzv. 'ice-pick' lobotomiju transorbitalnim pristupom koja se vršila i bez hospitalizacije,
a trajala je svega nekoliko minuta. Tako je samo u Americi, u razdoblju od 1939-1951.
godine, napravljeno više od 18000 lobotomija. Operacija je u to vrijeme predstavljala
'idealno rješenje' za sve oblike 'problematičnog ponašanja' (tako su se 'liječili'
čak i kronični bolni sindromi jer je nakon lobotomije nestajela zabrinutost
zbog boli). Npr. radila se zatvorenicima, rođacima koji su zbog raznoraznih
razloga proglašavani neuračunljivima ili nepodobnima (Rosemary Kennedy, sestra
američkog predsjednika Johna Kennedyja; Rose Williams, sestra američkog pisca
Tenessee Williamsa), pobunjenicima, političkim protivnicima (npr. holivudskoj
glumici Frances Farmer koja je simpatizirala komunizam), itd..
Raspon promjena ponašanja koje se vide u lobotomiranih bolesnika vrlo je velik.
Većina bolesnika izgube spontanost, sposobnost planiranja, procjene, odgode
nagrade (ugode), kajanja, apstraktnog mišljenja, kreativnosti, rješavanja problema,
mentalne fleksibilnosti (nastupa pretjerana konkretnost i vezanost za podražaj),
osjećaj jastva, sposobnost izvršavanja pravilnog redoslijeda radnji odnosno
praćenja instrukcija, donošenja strateških odluka temeljenih na pristupu različitim
rizicima, mogućnost istovremene pozornosti za više različitih događaja i mijenjanje
žarišta koncentracije, kapacitet usvajanja konteksta i složenih situacija, neotpornost
na distrakciju i interferenciju, inhibiciju neposrednih odgovora na podražaje,
itd.. Zbog navedenih učinaka, početkom pedesetih godina su se ovakve operacije
prestale izvoditi. Ipak, u nekim zemljama je za radikalnu kontrolu nasilnog
ponašanja lobotomija i danas dopuštena metoda.
Iako izraz 'frontalni sindrom' obuhvaća cijeli raspon navedenih promjena ponašanja,
svaki bolesnik može imati različitu raspodjelu i intenzitet izraženih deficita.
Specifični obrazac deficita u svakog pojedinog bolesnika određen je mjestom,
veličinom i prirodom oštećenja, kao i vremenskim tijekom oštećenja, te osobnošću
prije oštećenja.
U skladu sa slučajevima iz kliničke prakse, sindrom čeonog režnja može se grubo
podijeliti u dva tipa. U frontalnoj abuliji prevladava gubitak sposobnosti izvršavanja
voljnih radnji, nedostatak kreativnosti, znatiželje i inicijative, pa nastaje
apatija (nedostatak želja i emocionalna neosjetljivost). Odgovorno oštećenje
nalazi se većinom u dorzolateralnom dijelu čeonog režnja (tj. dijelovima čeonog
režnja koji se sastoje od heteromodalnog korteksa).
U sindromu frontalne disinhibicije bolesnik pokazuje dramatičnu impulzivnost,
zajedno s gubitkom sposobnosti prosuđivanja, uvida i predviđanja. Disocijacija
između relativno intaktnih 'intelektualnih spoznajnih' funkcija s jedne, i dramatično
promjenjenog načina ponašanja i emocionalnih reakcija s druge strane, može biti
vrlo izražena. Tako je, primjerice, opisan slučaj 63-godišnje žene koja je,
nakon operacije meningeoma u olfaktornom području, započela intimne odnose s
potpunim strancima, čak i s jednim koji je upravo izašao iz zatvora. Kad je
upitana zašto to čini, ona se složila da takvo ponašanje nije pametno, ali da
si ne može pomoći jer je 'izgubila kočnice'. Bolesnica nije promijenila ponašanje
čak ni kad joj je dotični ukrao njene stvari i prenio spolnu bolest. Prilikom
neuropsihološkog testiranja, tzv. 'frontalne kognitivne zadatke' rješavala je
normalno (npr. Lurijine motoričke slijedove, test sortiranja karata, itd.).
Postoperativne radiološke snimke pokazivale su da je operacijom oštećen samo
orbitofrontalni korteks, dok su dorzolateralna područja moždane kore bila intaktna.
U skladu s ovim i mnogim drugim slučajevima može se zaključiti da oštećenje
orbitofrontalnih i medijalnih čeonih areja (tj. dijelova čeonog režnja koji
se sastoje od paralimbičkog korteksa) dovode pretežno do sindroma disinhibicije.
Obrnutim slijedom, može se definirati orbitofrontalni korteks i druge paralimbičke
komponente čeonog režnja kao transmodalne čvorove za povezivanje misli, zapamćenih
sadržaja i iskustava s odgovarajućim emocionalnim i visceralnim stanjima. U
diferencijalnoj dijagnozi je najvažnije razlikovati abuliju od depresije,a sindrom
disinhibicije od manije.
'Radnim pamćenjem' nazivamo onu vrst pamćenja što je aktivna tek u kratkom vremenskom
razdoblju (obično nekoliko sekundi, npr. upamćivanje i nazivanje telefonskog
broja). Uloga neurona dorzolateralnog prefrontalnog korteksa u radnom pamćenju
otkrivena je snimanjem aktivnosti ovih neurona u majmuna prilikom testa odgođenog
odgovora. U testu majmun fiksira središnju točku monitora i uvježba se da tu
točku i dalje fiksira tijekom kratkotrajne pojave podražaja u nekom drugom dijelu
vidnog polja i razdoblja odgode nakon prikazivanja podražaja (3-5 sekundi).
Da bi dobio nagradu, tek potom majmun pogled usmjeri u onu točku vidnog polja
u kojoj se prethodno pojavio podražaj. Snimljena aktivnost prefrontalnih neurona
pokazuje da je ovakvo usmjeravanje pogleda majmuna vođeno isključivo kratkotrajnim
pamćenjem mjesta na kojem se podražaj pojavio. Ovi prefrontalni neuroni dakle
imaju 'memorijska polja', što se definiraju kao oni položaji podražaja u vidnom
polju na koje određeni neuron reagira najvećom frekvencijom okidanja, tj. kodira
samo tu prostornu točku vidnog polja. Nakon pojave i nestanka podražaja u svom
'memorijskom polju', takav neuron ostaje tonički aktivan tijekom razdoblja odgode,
a potom se njegova aktivnost naglo prekida. Štoviše, prilikom pojave podražaja
u dijametralno suprotnoj točci vidnog polja, aktivnost ovog neurona je inhibirana.
Budući da neuroni prefrontalnog korteksa s ovakvim svojstvima pokazuju selektivnu
aktivnost za vrijeme inhibicije odgovora u zadacima odgođenog odgovora, oni
vjerojatno igraju i ključnu ulogu u inhibiciji impulsa koji nisu prikladni za
kontekst. Proširujući navedene rezultate može se pretpostaviti da se posredovanjem
radnog pamćenja reprezentacije mnogobrojnih vanjskih događaja i internih fenomena
mogu istovremeno i interaktivno procesirati na način da se središte pozornosti
mijenja od jednog do drugog. No, radno pamćenje nije uključeno samo u prostornu
pozornost kao što je to ovdje ilustrirano, nego i u sve druge vidove kognitivne
i lingvističke obrade informacija. Stoga se, poopćavajući navedeno, može zaključiti
da je radno pamćenje odraz sposobnosti moždane kore da u svijest prizove upamćene
sadržaje i u odsutnosti podražaja (apstraktno mišljenje), što je vjerojatno
jedan od najvažnijih evolucijskih napredaka moždanog ustrojstva, jer ono pomoću
isključivo mentalne manipulacije omogućuje povezanost i kontinuitet prošlih
iskustava i trenutnih djelovanja. Mogućnost 'vrtenja' alternativnih scenarija
događaja promovira kontekstualnu fleksibilnost, predviđanje i planiranje radnji
s obzirom na prostor i vrijeme.
Mnogi neuroni čeonog režnja koji se aktiviraju vidnim podražajima imaju manju
specifičnost za boju, veličinu ili orijentaciju podražaja, nego za njegovu biološku
važnost. Naime, neki prefrontalni neuroni koji živahno odgovaraju na podražaj
povezan s nagradom mogu promijeniti svoj odgovor kad isti podražaj bude povezan
s neugodnim ili neutralnim ishodom. Ovakvi odgovori neurona prefrontalnog korteksa
vjerojatno pomažu uspostavljanju subjektivne realnosti koja je osjetljiva na
važnost podražaja, a ne na njegova površinska svojstva.
S obzirom na strukturu, veze i funkcije, lako se uočava da je korteks čeonog
režnja čovjeka toliko heterogen i složen da ni jedan pojedinačni opis ne može
objasniti sve njegove uloge u ponašanju. U pokušaju da se to ipak učini, prefrontalni
korteks može se konceptualizirati kao mjesto na kojem konfluiraju dvije goleme
mreže: jedna za radno pamćenje, izvršne funkcije i pozornost, tj. kontrolu djelovanja
(s transmodalnim epicentrima u prefrontalnom i stražnjem parijetalnom korteksu),
i druga za kontrolu ponašanja (s transmodalnim epicentrima u prefrontalnom korteksu
i orbitofrontalnom paralimbičkom korteksu). Mediodorzalna jezgra talamusa i
glava kaudatusa su dodatne neophodne komponente obiju mreža.
Budući da zbog ostvarivanja ponašanja prikladnog kontekstu prefrontalni korteks
orkestrira svim drugim kortikalnim mrežama i međumrežnim interakcijama, njegova
ozljeda ne rezultira očiglednim deficitima kao što je to slučaj u drugim mrežama,
već deficitom u nadzoru nad ponašanjem koje nije apsolutno već zavisno od situacije.
Stoga se klinički uočljivi deficiti obično vide tek nakon masivnog obostranog
oštećenja, a samo katkad i nakon opsežnijeg jednostranog oštećenja (promjene
ponašanja mogu biti tako suptilne da su uočljive samo osobama koje dobro poznaju
osobnost bolesnika prije oštećenja). I ovdje se uočava lateralizacija kortikalnih
funkcija, jer nakon oštećenja lijeve hemisfere može nastati sindrom abulije,
dok jednostrano oštećenje desne hemisfere može prouzročiti disinhibirano ponašanje.
Mnogobrojne su teorije koje povezuju oštećenje razvitka mreža prefrontalnog
korteksa s nastankom autističnih poremećaja u djece: autizma, Aspergerovog sindroma
(po definiciji slabije izraženog oblika autizma kod kojeg su kognitivne funkcije
manje oštećene, a govor nije poremećen) i 'high-functioning' autizma (najblaži
oblik). Iako je raspon autističnih poremećaja velik, sve ih u manjoj ili većoj
mjeri karakteriziraju otežana socijalna interakcija i komunikacijske sposobnosti,
ponavljajući obrasci ponašanja i ograničen interesi. Otežana socijalna interakcija
u autista se odražava kroz repetitivni, stereotipni ili idiosinkratični govor,
nedostatak empatije tj. gubitak anticipacije za emocije, intencije i djelovanje
drugih, ali i vlastito djelovanje (ali s neoštećenom sposobnošću za izvršavanje
pokreta), ograničeni repertoar ponašanja, deficit u mijenjanju ciljeva od lokalnog
prema globalnom procesiranju. Jedan od najkarakterističnijih znakova pri utvrđivanju
autističnog poremećaja u djece je nesposobnost za zamjenu uloga u igri (poremećeno
doživljavanje vlastitog ja), odnosno nemogućnost transformacije objekata (npr.
za razliku od normalnog trogodišnjaka, autistično dijete neće uzeti bananu u
ruku i pomoću nje 'telefonirati' ili sjesti naopačke na stolac i 'voziti formulu',
niti shvatiti što bi ono tu trebalo napraviti). Činjenica da mnogi od ovih bolesnika
imaju iznadprosječnu sposobnost koncentracije i usmjeravanja pažnje na jedno
specifično područje od interesa dodatno govori u prilog povezanosti oštećenja
prefrontalnih mreža i njihovog ponašanja. Kako abnormalnosti multimodalne integracije
dovode do sklonosti lokalnim podražajima, to je procesiranje više specijalizirano
za elementarna fizička obilježja podražaja. Primjerice, opisan je autistični
dječak koji bi po razbacivanju šibica na pod, u tren oka nepogrešivo ustanovljavao
njihov točan broj, npr. 141! Slični su slučajevi dokumentirani i za druge modalitete.
Neuropatološki su opisane razne promjene od kojih je možda najvažnija abnormalnost
minikolumnarizacije prefrontalnog (naročito BA9) i lateralnog temporalnog korteksa
(BA21 i 22).
5. Završne napomene i zaključci
U odnosu na druge životinjske vrste,
slijedeća su obilježja ponašanja jedinstvena za čovjeka: lokomocija se temelji
na nogama, tako da su ruke oslobođene za druge složene funkcije; palac je odmaknut,
a fina motorike šake omogućuje razvitak oruđa i tehnologija; govor i jezik podloga
su naprednih oblika komunikacije i simboličkog mišljenja: razvijene socijalne
interakcije i kultura (što su dovele do podjele posla u društvu, te kontrole
seksualnih nagona i agresije); produljeno razdoblje odrastanja; individualni
umjetnički i duhovni izričaj. Sva navedena obilježja izravno ovise o moždanoj
kori, pa se i neurobiologija ponašanja čovjeka u najvećem dijelu odnosi na proučavanje
moždane kore.
Od epohalnog otkrića da je lijeva hemisfera mozga čovjeka superiorna za jezične
funkcije što ga je načinio Broca 1865. godine, otkrivene su i brojne druge strukturalne
i funkcionalne asimetrije mozga čovjeka. Svrha asimetrije nije potpuno jasna,
ali vjerojatno predstavlja odraz biološke prednosti u smislu koncentriranja
kontrole pojedinih dijelova mreža u istoj hemisferi, s ciljem minimaliziranja
zaostajanja u procesiranju uslijed prijenosa informacija u suprotnu hemisferu.
Mnogi osjetni doživljaji koji inače aktiviraju asocijativna i transmodalna područja
moždane kore nisu dostupni svijesti, dakle utječu na ponašanje iako ih čovjek
nije svjesno doživio. Neki dobro poznati i metodama slikovnog prikaza aktivnosti
mozga proučeni primjeri su fenomen rezidualnog vida u hemianopsičnih bolesnika,
implicitno pamćenje i 'priming' u bolesnika s globalnom amnezijom, odgovori
autonomnog živčanog sustava na poznata lica u prozopagnozičara, nesvjesna percepcija
feromona i ideomotorno djelovanje.
S obzirom na zadivljujuću složenost moždane kore čovjeka, naročito one čeonog
režnja, ne iznenađuje činjenica da različite bolesti, poremećaji i ozljede što
zahvaćaju moždanu koru mogu dovesti do iznimno složene simptomatologije. Iako
udžbenici neuroznanosti često teže naglašavanju čvrstih i nepromjenjivih odnosa
između strukture i funkcije (npr. oštećenje optičkog trakta uvijek dovodi do
kontralateralne homonimne hemianopsije), ovakav pristup ponašanju i kogniciji
nije zadovoljavajući jer strukturni temelj ovih procesa ima oblik velikih preklapajućih
neuronskih mreža organiziranih oko recipročno povezanih kortikalnih epicentara.
Pojedini dijelovi ovih mreža mogu se podijeliti na ključna i participirajuća
područja. Oštećenja koja nepovratno dovode do nesposobnosti djelovanja u određenoj
domeni ponašanja ili kognicije pomažu nam odrediti dijelove mreže koji su neophodni
za integritet određene kognitivne domene, a tek aktivacije pojedinih dijelova
mozga snimljene funkcionalnim tehnikama slikovnog prikaza u trenutku izvršavanja
zadataka povezanih s dotičnom domenom otkrivaju koja još područja mozga sudjeluju
u njenom koordiniranom djelovanju. Stoga za dobivanje što potpunije slike odnosa
između strukture mozga i ponašanja, treba tradicionalni pristup utemeljen na
proučavanju pacijenata s žarišnim oštećenjem mozga povezati s eksperimentalnim
podacima dobivenim tehnikama slikovnog prikaza aktivacije mozga (fMRI, PET).